
Par Patrick Pla, Université Paris-Saclay
Les vésicules extracellulaires (VE) sont des particules produites et excrétées par les cellules et entourées d’une bicouche lipidique. Elles étaient autrefois considérées comme des déchets cellulaires mais elles constituent en fait une forme importante de communication intercellulaire et attirent désormais une attention considérable en raison de leurs rôles dans une variété de processus physiologiques et pathologiques.
- Production des vésicules extracellulaires
- Effets des vésicules extracellulaires sur les cellules-cibles
Production des vésicules extracellulaires
L’une des principales classes de VE est constituée par les exosomes, de petites vésicules, dont la taille varie de 30 à 150 nm de diamètre, qui se forment dans des corps multivésiculaires (MVB) et sont acheminées vers la surface cellulaire (au lieu des lysosomes où ces corps multivésiculaires aboutissent habituellement). Les MVB fusionnent avec la membrane plasmique et libèrent leur contenu, les exosomes, dans l’espace extracellulaire. Un tel processus a pu être observé in vivo par exemple lors de la formation du réseau des trachées chez la drosophile où des exosomes sont émis dans la lumière trachéale (Camelo et al., 2022).
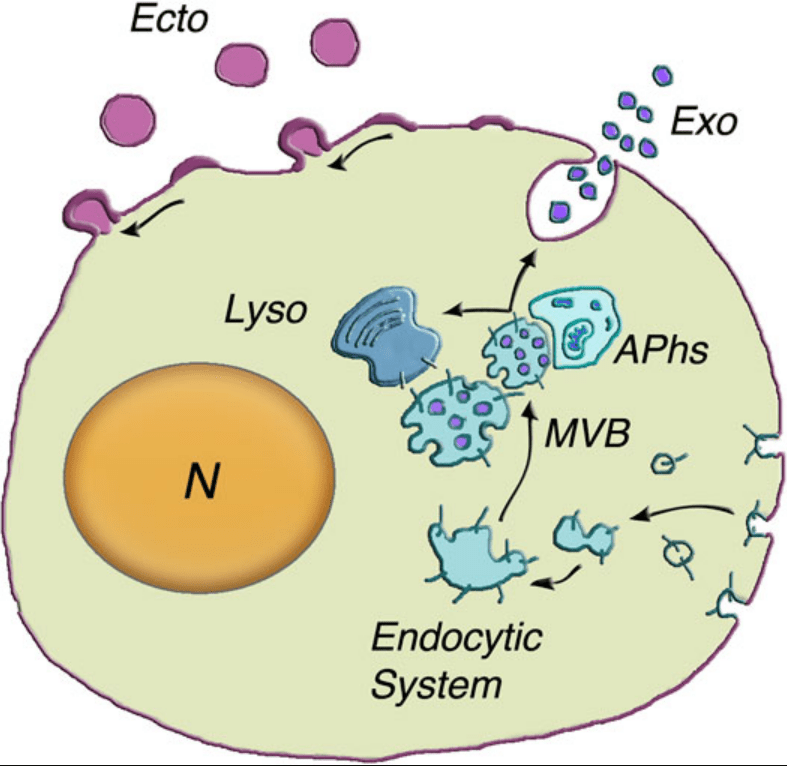
L’autre classe majeure de VE est le plus souvent appelée microvésicules (MV). Les MV sont nettement plus grandes que les VE dérivées des MVB et leur diamètre varie de 0,2 à 1,0 μm de diamètre et se forment à la suite de leur bourgeonnement vers l’extérieur et de leur fission de la membrane plasmique (Desrochers et al., 2016; Latifkar et al., 2019).
Les VE peuvent voyager d’une cellule productrice à une cellule réceptrice dans le milieu extracellulaire des tissus ou être transportées par le sang ou le liquide céphalo-rachidien. On en trouve également dans les urines, la salive, les larmes (sans que cela ait forcément une valeur adaptative). Des nouvelles techniques de protéomiques quantitatives permettent de trouver l’origine cellulaire des VE dans les biofluides selon leur profil protéique, notamment en tétraspanines dont les qualités/quantités varient beaucoup selon le type cellulaire d’origine et sont donc des marqueurs intéressants (Garcia-Martin et al., 2022).
Une variété de protéines, d’ARNm et de micro-ARN est associée aux MV et aux exosomes. Cette cargaison est souvent localisée dans la lumière des VE, même si, dans certains cas, elle est associée à la surface de la vésicule (Jeppesen et al., 2019; Valadi et al., 2007; Desrochers et al., 2016; Ratajczak et al., 2006).
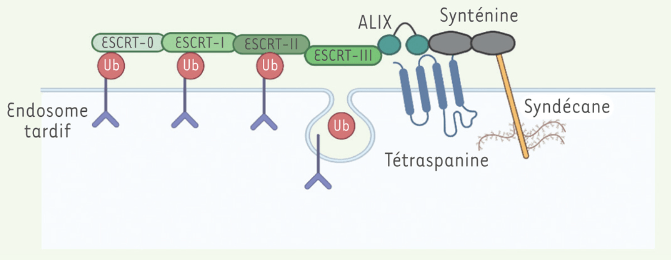
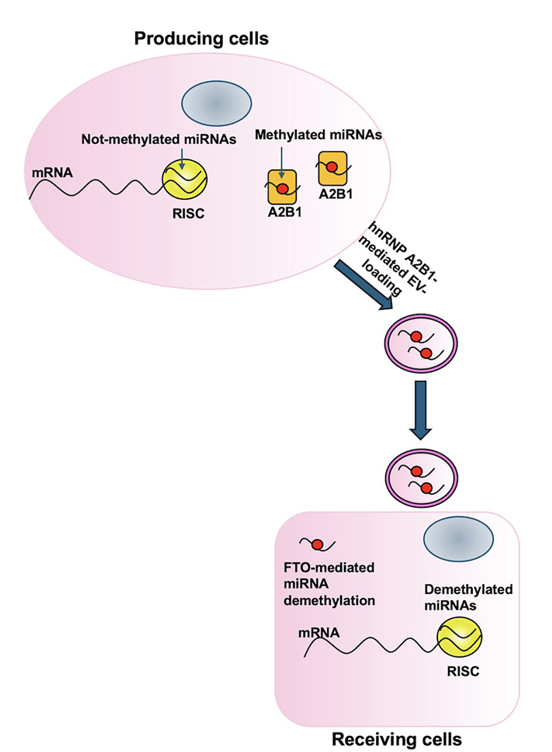
Plusieurs protéines de liaison à l’ARN (RBP) ont été impliquées dans la sélection et le chargement des ARN dans les VE, notamment les protéines nucléaires hétérogènes ribonucléoprotéines A2/B1 (hnRNPA2B1) (Villarroya-Beltri et al., 2013), SYNCRIP (Santangelo et al., 2016), HuR (Mukherjee et al., 2016) et les protéines MVP (Statello et al., 2018; Teng et al., 2017). YBX1 a été impliqué dans le chargement du microARN miR-233 dans les exosomes (Shurtleff et al., 2016).
La sélection des microARN qui seront chargés dans les vésicules extracellulaires peut se faire par des modifications de leurs nucléotides comme par exemple la méthylation du carbone 6 de l’adénosine (N6-methyladenosine (m6A)) catalysée par la methyltransferase-like 3 (METTL3) (Garbo et al., 2024). Cette méthylation rend les microARN inactifs. La méthylation est réversible et la déméthylation qui se déroule dans les cellules receveuses permet aux microARN d’y inhiber la traduction de leur ARNm cibles.
La réception des vésicules extracellulaires par les cellules-cibles commence à être étudiée en détails. Des mécanismes comme la fusion directe avec la membrane plasmique, l’endocytose et la phagocytose ont été observés (Mulcahy et al., 2014).
Effets des vésicules extracellulaires sur les cellules-cibles
Comme nous venons de le voir dans la figure précédente, les MV et les exosomes peuvent transférer leur cargaison vers d’autres cellules (c’est-à-dire receveuses) et, ce faisant, modifier le comportement cellulaire. La réception des vésicules extracellulaires par les cellules-cibles commence à être étudiée en détails. Des mécanismes comme la fusion directe avec la membrane plasmique, l’endocytose et la phagocytose ont été observés (Mulcahy et al., 2014).
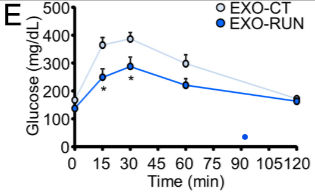
Lors d’un exercice musculaire intense les cellules musculaires striées squelettiques relarguent des exosomes contenant des microARN à destination du foie qui aboutissent à une meilleure sensibilité à l’insuline et donc à une meilleure régulation de la glycémie (Castano et al., 2020).
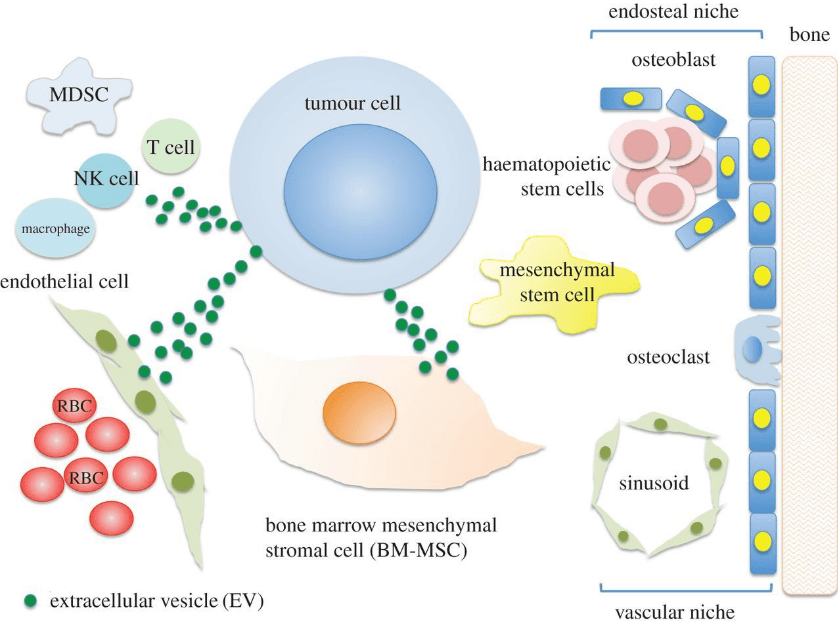
Les rôles des VE dans la progression du cancer ont été étudiés. Elles aident à façonner le microenvironnement tumoral, à promouvoir l’immunosuppression et à améliorer la croissance, la survie, l’invasion et la propagation métastatique des cellules cancéreuses (Antonyak et Cerione, 2014; Antonyak et al. ., 2011; Costa-Silva et al., 2015; Kreger et al., 2016; Van Niel et al., 2018). Par exemple, des cellules faiblement métastatiques issues de cancer du sein secrètent des vésicules extracellulaires riches en Transglutaminase 2 (Tg2) qui activent les fibroblastes environnants à produire une matrice extracellulaire favorable à la migration (Schwager et al., 2022). Les VE ont été particulièrement bien étudiées dans le cas des tumeurs des cellules sanguines dans la moelle osseuse (Ohyashiki et al., 2017).
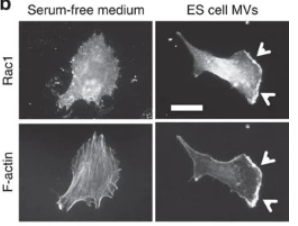
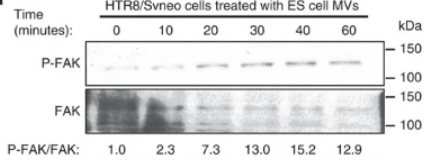
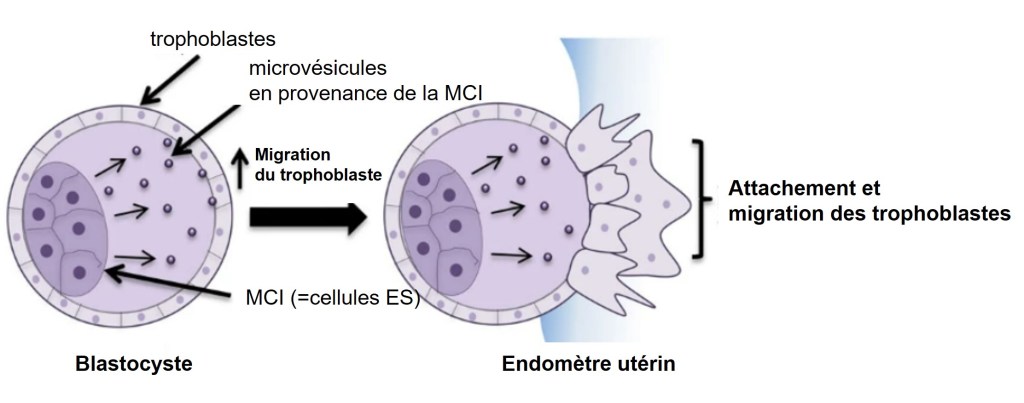
Cependant, les VE ont également un impact sur un large éventail de processus physiologiques. Dans les blastocystes de souris, Les VE produites par les cellules pluripotentes dans l’ICM sont transférés au trophectoderme pour favoriser l’implantation, une étape précoce de la grossesse dans laquelle l’embryon en développement se fixe et envahit l’utérus (Desrochers et al., 2016). Les MV transportent alors la laminine et fibronectine qui interagissent avec les intégrines le long des surfaces des trophoblastes, et déclenchent l’activation de deux kinases de signalisation, JNK et FAK, ce qui stimule la migration des trophoblastes.
Dans le système nerveux, de nombreuses fonctions des vésicules extracellulaires ont pu être mises en évidence à la fois en situation développementale, physiologique et pathologique. Par exemple, au cours du développement, les astrocytes produisent des VE qui exposent à leur surface la protéine synapsine-1 qui se lie au récepteur neuronal NCAM (neuronal cell adhesion molecule). L’internalisation de cette protéine après sa fixation à son récepteur par les neurones favorise la survie neuronale et la croissance des dendrites et des axones (Wang et al., 2011).

Les exosomes peuvent avoir un rôle dans la régulation des cellules souches et dans le vieillissement. Par exemple, des exosomes produits par des adipocytes péri-musculaires de souris âgées (mais pas de souris jeunes) contiennent de grandes quantités de miARN Let-7d-3p qui inhibent la prolifération des cellules souches musculaires (les cellules satellites) et des précurseurs musculaires en diminuant l’expression du facteur de transcription HMGA2 (Itokazu et al., 2022).
EN DIRECT DES LABOS :
- Adhérences cellule-cellule
- Arabidopsis thaliana
- Axe antéro-postérieur chez la drosophile
- Biomécanique du développement
- Caenorhabditis elegans
- Concepts principaux
- Contrôle de la traduction
- Contrôle de la transcription
- Contrôle génétique et épigénétique
- Croissance du tube pollinique et double fécondation chez les Angiospermes
- Croissance et guidage axonal
- Des modèles animaux moins classiques
- Développement de l’oeil des Vertébrés
- Développement et évolution
- Et l’Humain ?
- Exercices sur l’ovogenèse, la spermatogenèse et la fécondation
- Exercices sur le contrôle de l’expression des gènes
- Exercices sur le développement des bourgeons de membre
- Exercices sur le développement des muscles striés squelettiques
- Exercices sur le développement des végétaux et les hormones végétales
- Exercices sur les cycles et les divisions cellulaires
- Exercices sur les étapes du développement, les inductions embryonnaires et la mise en place des axes de polarité
- Exercices sur les matrices extracellulaires, le cytosquelette et les adhérences cellule-cellule
- Exercices sur les voies de signalisation
- Glossaire
- Glossaire des termes liés à la génétique
- Glossaire des termes liés au cytosquelette, la matrice extracellulaire, l’adhérence et la migration cellulaire
- Hématopoïèse et développement des cellules du système immunitaire
- Histoire de la biologie cellulaire et de la biologie du développement
- L’acide rétinoïque
- L’apoptose
- L’autophagie
- L’organogenèse
- L’ovogénèse prépare le développement embryonnaire
- La drosophile
- La fécondation
- La formation des somites
- La gastrulation
- La gastrulation (version allégée)
- La métamorphose chez les Hexapodes et les Amphibiens
- La neurogénèse chez les mammifères adultes
- La neurulation
- La poule
- La signalisation calcique
- La souris
- La superfamille TGFβ et ses voies de signalisation
- La voie de signalisation de l’auxine et ses rôles
- La voie de signalisation Hedgehog
- La voie de signalisation Hippo et ses composants YAP/TAZ
- La voie de signalisation Notch
- Le clivage
- Le cytosquelette
- Le destin des cellules et les réseaux de régulation génique
- Le développement des bourgeons de membre
- Le développement des muscles striés squelettiques
- Le développement des organes génitaux et des cellules germinales
- Le développement du cortex
- Le méristème apical caulinaire en phase végétative et lors de la formation d’une fleur
- Le poisson zèbre
- Le xénope
- Les cellules des crêtes neurales
- Les cellules et les gènes en action dans le développement
- Les cellules souches
- Les cellules tumorales
- Les cycles et les divisions cellulaires
- Les étapes du développement
- Les étapes du développement embryonnaire d’Arabidopsis thaliana et leur contrôle
- Les inductions embryonnaires et les gradients de morphogène
- Les matrices extracellulaires animales
- Les organismes modèles
- Les outils pour étudier l’expression et la fonction des gènes
- Les parois des cellules végétales
- Les techniques et les outils pour la biologie cellulaire
- Les transitions épithélio-mésenchymateuses et les migrations cellulaires
- Les voies de signalisation
- Les voies de signalisation FGF
- Mise en place des axes chez les Vertébrés
- Structures et processus cellulaires
- Voies de signalisation WNT
