

Arabidopsis thaliana est devenu depuis les années 1990, le modèle principal d’étude du développement et la génétique des plantes (son utilisation en tant que modèle a été initiée par le botaniste allemand Friedrich Laibach en 1907). C’est un paradoxe car cette plante n’a aucune valeur agricole mais elle possède de nombreux avantages.
Arabidopsis thaliana est une Brassicacée annuelle qui a une distribution étendue à travers toute l’Europe et l’Amérique du Nord et une bonne partie de l’Asie. Son nom commun est l’arabette des dames ou arabette de Thalius.
Sa petite taille permet de cultiver de très nombreux plants sur une surface restreinte.
Son cycle de vie dure seulement 6 semaines. Un plant peut produire jusqu’à 5000 graines, ce qui permet de faire de la génétique sur une vaste population. La petite taille de la plante (hauteur adulte de 15 à 25 cm) permet la culture de plusieurs dizaines d’individus par mètre carré, ce qui permet de réaliser des analyses poussées sur des populations sans que cela prenne trop de place.
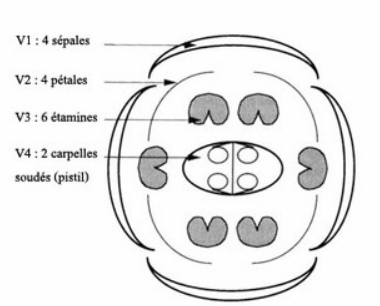
Les fleurs apparaissent environ 3 à 4 semaines après la germination. Chaque fleur possède 4 sépales et 4 pétales, disposés en croix (d’où l’ancien nom de Crucifère à la famille des Brassicacées), avec un décalage de 45° entre les sépales et les pétales. Plus à l’intérieur, on trouve 6 étamines et 2 carpelles qui contiennent des ovules.

Les gamètes sont formés dans des organismes particuliers appelés gamétophytes. Le gamétophyte mâle est le grain de pollen et le gamétophyte femelle est le sac embryonnaire qui ne sort pas de l’ovule.



On peut réaliser des auto-fécondations chez ce qui facilite les études génétiques par l’obtention simple de mutants homozygotes. En effet, les gènes SKR et SCR qui limitent l’auto-pollinisation chez d’autres plantes présentent des mutations perte-de-fonction chez Arabidopsis thaliana. Un eespèce proche, Arabidopsis lyrata possède, en revanche des gènes SKR et SCR fonctionnels et leur expression dans Arabidopsis thaliana inhibe sa capacité d’autofécondation (Strickler et al., 2013).
Entre la pollinisation et la fécondation, les tubes polliniques se développent de manière invasive dans le pistil et permettent aux deux spermatozoïdes de pénétrer dans le sac embryonnaire intégré dans l’ovule. Le sac embryonnaire est composé de deux synergides, une oosphère (=gamète), une cellule centrale (à deux noyax haploïdes) et trois cellules antipodales (Yang et al., 2010). Les tissus femelles, notamment les ovules, émettent des signaux pour guider les tubes polliniques dans le sac embryonnaire (Higashiyama et al., 2016). Les synergides sécrètent de petites molécules sous l’induction non cellulaire autonome des cellules centrales pour attirer les tubes polliniques. Les signaux peptidiques sont perçus par les récepteurs à activité kinase exprimées dans les tubes polliniques, puis transduites vers des machineries de croissance intracellulaire telles que de petites GTPases et le cytosquelette, qui sont régulées par un gradient de Ca2+ (Krichevsky et al., 2007; Steinhorst et al., 2013).
Il y a une double fécondation : un spermatozoïde féconde l’oosphère ce qui donne le zygote qui donne l’embryon et un autre spermatozoïde (provenant du même grain de pollen que le précédent) féconde la cellule centrale ce qui est à l’origine de l’albumen, tissu de réserve triploïde (car la cellule centrale contenait deux noyaux haploïdes).
Après la fécondation, le début du développement embryonnaire d’Arabidopsis comprend une succession de divisions très reproductibles, ce qui permet de déceler facilement la survenue de toutes différences chez les mutants. Des cartes des territoires présomptifs sont disponibles permettant de connaître le destin des différentes cellules.
L’ovule met 2 semaines à se transformer en graine.
Des tissus ou des cellules particulières d’A. thaliana peut être particulièrement étudiés et devenir en eux-mêmes des modèles. Citons les trichomes. Ce sont des cellules épidermiques uniques que l’on trouve sur les feuilles en rosette et sur la tige. Le développement des trichomes est principalement étudié sur les feuilles en rosette, où ils sont espacés régulièrement. Les trichomes en différenciation subissent typiquement quatre cycles d’endoréduplication et forment trois ou quatre branches. La recherche systématique de mutants de trichomes a révélé un grand nombre de mutants interférant avec divers processus de développement tels que la formation de motifs, la régulation du cycle cellulaire et la morphogenèse cellulaire.

Le génome assez compact d’Arabidopsis thaliana (135 millions de pb répartis sur 5 chromosomes, ce qui est très petit pour un Angiosperme) a été entièrement séquencé en l’an 2000. Il possède 29.000 gènes ce qui est du même ordre de grandeur que le génome de la souris ou de l’humain. Il y a 225 gènes par mégabases chez Arabidopsis contre 7 chez Homo sapiens. Contrairement à beaucoup d’espèces cultivées qui présentent des polyploïdies complexes, Arabidopsis reste majoritairement diploïde, ce qui simplifie les analyses génétiques, notamment l’identification des mutations récessives. Cependant, de larges portions du génome ont été dupliquées au cours de son histoire évolutive et il existe de nombreuses familles multigéniques.
On peut assez facilement créer des plants transgéniques en infectant des cellules avec une forme génétiquement modifiée de la bactérie Agrobacterium tumefaciens.
Les propriétés de la bactérie pathogène Agrobacterium tumefaciens (qui provoque habituellement des galles) sont utilisées comme vecteur des gènes. On utilise le plus souvent des plasmides dérivés de son plasmide Ti dont une partie, l’ADN-T est habituellement transféré dans le génome de la plante. Pour la transgénèse, on utilise des plasmides Ti désarmés (ou D-Ti), car leur ADN ne porte plus les gènes responsables du pouvoir pathogène, et l’ADN-T contient les gènes que l’on souhaite étudier et un ou plusieurs gènes de sélection (le gène de résistance à la kanamycine par exemple). L’ADN-T modifié s’insère dans le génome et des agents de sélection permettent de ne faire survivre que les cellules végétales transformées. Des plantes entières sont ensuite régénérées à partir des protoplastes (cellules végétales sans parois) ou des cals où la transformation a eu lieu par des techniques classiques de culture in vitro, faisant intervenir les hormones végétales telles que l’auxine et les cytokinines.
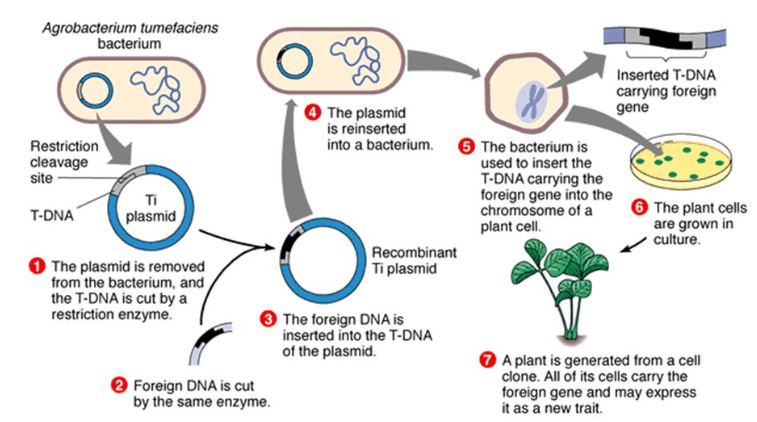
Les chercheurs peuvent aussi utiliser le fait que l’ADN-T d’Agrobacterium s’insère au hasard dans le génome et ils peuvent étudier les conséquences phénotypiques de ces mutations insertionnelles qui peuvent casser une séquence codante ou régulatrice d’un gène.
Malgré sa domination claire dans les publications de génétique et de biologie cellulaire du développement des plantes, il est clair qu’Arabidopsis ne constitue qu’un modèle particulier. Certains phénomènes ne peuvent être étudiés chez elle comme la formation du bois ou des feuilles composées. Cela dit, les chercheurs ont réussi à produire un équivalent de bois (du xylème secondaire) chez Arabidopsis en lui ajoutant des poids, indiquant que le poids porté par la tige déclenche le développement des tissus secondaires et que ce programme était toujours activable chez Arabidopsis (Ko et al., 2004).
POUR ALLER PLUS LOIN :
Article de rétrospective de 50 années de recherche sur Arabidopsis.
QUELQUES LABORATOIRES FRANCOPHONES QUI TRAVAILLENT SUR CE MODELE :
Equipe « Génome et développement des plantes » – Université de Perpignan
Equipe « Hormones, nutriments et développement » – Institut des Sciences des Plantes de Montpellier
Equipe « Morphogenèse florale » – Université de Lyon
- Adhérences cellule-cellule
- Axe antéro-postérieur chez la drosophile
- Biomécanique du développement
- Caenorhabditis elegans
- Concepts principaux
- Contrôle de la traduction
- Contrôle de la transcription
- Contrôle génétique et épigénétique
- Croissance du tube pollinique et double fécondation chez les Angiospermes
- Croissance et guidage axonal
- Des modèles animaux moins classiques
- Développement de l’oeil des Vertébrés
- Développement et évolution
- Et l’Humain ?
- Exercices sur l’ovogenèse, la spermatogenèse et la fécondation
- Exercices sur le contrôle de l’expression des gènes
- Exercices sur le développement des bourgeons de membre
- Exercices sur le développement des muscles striés squelettiques
- Exercices sur le développement des végétaux et les hormones végétales
- Exercices sur les cycles et les divisions cellulaires
- Exercices sur les étapes du développement, les inductions embryonnaires et la mise en place des axes de polarité
- Exercices sur les matrices extracellulaires, le cytosquelette et les adhérences cellule-cellule
- Exercices sur les voies de signalisation
- Glossaire
- Glossaire des termes liés à la génétique
- Glossaire des termes liés au cytosquelette, la matrice extracellulaire, l’adhérence et la migration cellulaire
- Hématopoïèse et développement des cellules du système immunitaire
- Histoire de la biologie cellulaire et de la biologie du développement
- L’acide rétinoïque
- L’apoptose
- L’autophagie
- L’organogenèse
- L’ovogénèse prépare le développement embryonnaire
- La drosophile
- La fécondation
- La formation des somites
- La gastrulation
- La gastrulation (version allégée)
- La métamorphose chez les Hexapodes et les Amphibiens
- La neurogénèse chez les mammifères adultes
- La neurulation
- La poule
- La signalisation calcique
- La souris
- La superfamille TGFβ et ses voies de signalisation
- La voie de signalisation de l’auxine et ses rôles
- La voie de signalisation Hedgehog
- La voie de signalisation Hippo et ses composants YAP/TAZ
- La voie de signalisation Notch
- Le clivage
- Le cytosquelette
- Le destin des cellules et les réseaux de régulation génique
- Le développement des bourgeons de membre
- Le développement des muscles striés squelettiques
- Le développement des organes génitaux et des cellules germinales
- Le développement du cortex
- Le méristème apical caulinaire en phase végétative et lors de la formation d’une fleur
- Le poisson zèbre
- Le xénope
- Les cellules des crêtes neurales
- Les cellules et les gènes en action dans le développement
- Les cellules souches
- Les cellules tumorales
- Les cycles et les divisions cellulaires
- Les étapes du développement
- Les étapes du développement embryonnaire d’Arabidopsis thaliana et leur contrôle
- Les inductions embryonnaires et les gradients de morphogène
- Les matrices extracellulaires animales
- Les organismes modèles
- Les outils pour étudier l’expression et la fonction des gènes
- Les parois des cellules végétales
- Les techniques et les outils pour la biologie cellulaire
- Les transitions épithélio-mésenchymateuses et les migrations cellulaires
- Les vésicules extracellulaires
- Les voies de signalisation
- Les voies de signalisation FGF
- Mise en place des axes chez les Vertébrés
- Structures et processus cellulaires
- Voies de signalisation WNT
