
Par Patrick Pla, Université Paris-Saclay
SOMMAIRE :
La rencontre des gamètes
Contact et fusion entre les gamètes
Conséquences de la fécondation
La procréation médicalement assistée
Notre corps a un temps de vie limité mais la reproduction permet à l’espèce de se perpétuer. Ce sont les cellules de la lignée germinale (les gamètes) qui réalisent la reproduction sexuée. La fécondation correspond à la rencontre entre le gamète mâle (spermatozoïde) et le gamète femelle (ovocyte). Il s’agit de deux cellules très différentes : l’une est très mobile, l’autre l’est peu et son cytoplasme formera le cytoplasme du zygote issu de la fusion des deux cellules. Pour souligner cette différence, on parle d’anisogamie. Des mécanismes de reconnaissance et d’activation mutuelle permettent à ces cellules d’interagir, de fusionner et de mettre en commun leur matériel génétique. Les gamètes sont des cellules haploïdes ; leur fusion permet de rétablir la diploïdie habituelle des animaux.
Des dysfonctionnements dans la reproduction touchent 1 couple sur 10 en France et des solutions thérapeutiques d’assistance médicale à la procréation se sont développées à partir du début des années 1980s.
La rencontre des gamètes
La fécondation est externe chez les anoures tels que le xénope, avec néanmoins un accouplement qui s’appelle un amplexus. Le mâle maintient la femelle avec ses membres antérieurs et émet ses spermatozoïdes en même temps que les ovocytes sont émis du cloaque de la femelle. Cela permet de concentrer sur un espace limité et dans le temps la présence des gamètes mâles et femelles, ce qui optimise les rencontres entre les deux et limite les pertes.
Les spermatozoïdes entrent en contact avec la gangue gélatineuse qui entoure la masse des ovocytes et cette interaction est nécessaire au bon déroulement de la fécondation.
Chez les Mammifères, après l’éjaculation, les spermatozoïdes passent le col de l’utérus en traversant la glaire cervicale dont le passage est facilité autour de la période d’ovulation sous le contrôle du pic d’œstrogènes (mailles plus larges, pH moins acide). Chez l’Homme, seuls 2 millions de spermatozoïdes sur les 60 millions éjaculés traversent cette barrière. L’observation des modifications de la glaire cervicale au cours du cycle sexuel féminin est à la base d’une méthode de contraception (la méthode de Billings) dont l’efficacité est cependant très inférieure à la pilule contraceptive et au préservatif.
La nage flagellaire n’est pas le paramètre principal qui explique le déplacement des spermatozoïdes (on trouve des spermatozoïdes dans l’oviducte 30 min après éjaculation, ce qui est trop court pour que ce soit la seule mobilité flagellaire qui soit impliquée). Ce sont les contractions vaginales et utérines (contrôlées par les muscles lisses) qui les amènent vers l’oviducte. Ces contractions sont sous le contrôle de l’ocytocine dont la sécrétion est stimulée par l’accouplement. Les contractions utérines sont facilitées par les œstrogènes dont la concentration sanguine est maximale autour de la période de l’ovulation.
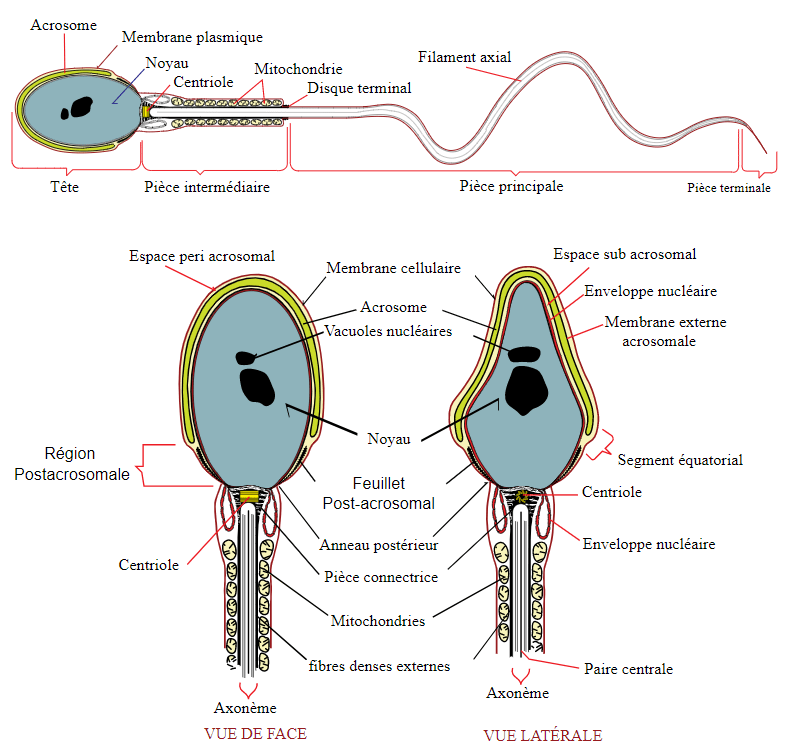
Un spermatozoïde éjaculé ne pourra pas directement féconder un ovocyte. Des modifications induites par les voies génitales femelles (utérus et surtout oviducte) doivent avoir lieu. C’est la capacitation. Les propriétés membranaires sont alors modifiées avec une perte de cholestérol membranaire et une concentration à l’avant de l’acrosome des radeaux lipidiques membranaires importants pour la fécondation. Il y a hyperactivation de la nage liée à une entrée de Ca2+ dans le cytoplasme qui aboutit à la phosphorylation et à l’activation des dynéines flagellaires. Des récepteurs spermatiques sont démasqués (perte des glycosides de surface ajoutés dans l’épididyme qui bloquent les récepteurs aux protéines de la zone pellucide de l’ovocyte). Les microfilaments d’actine sont réorganisés ce qui est essentiel pour la future réaction acrosomiale (Schiavi-Ehrenhaus et al., 2022). Tous ces phénomènes sont post-traductionnels car il n’y a plus de transcription, ni de traduction dans les spermatozoïdes matures.
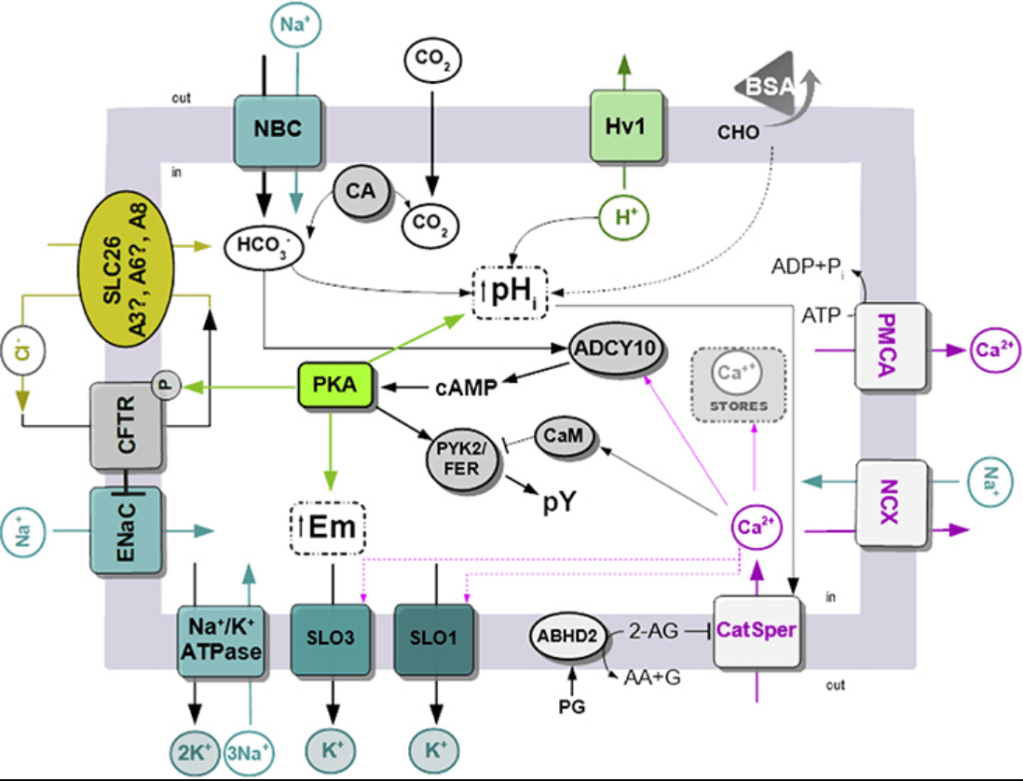
C’est au sein de l’oviducte que la nage flagellaire devient cruciale. Les spermatozoïdes ont besoin de mécanismes de navigation pour nager dans la bonne direction. Ces mécanismes de navigation reposent sur des signaux biochimiques et biophysiques externes (Tung et al., 2015). Les spermatozoïdes sont guidés par un gradient de température (thermocline de 2°C) le long de l’oviducte (de l’entrée à la région appelée ampoule où a lieu la fécondation). Les spermatozoïdes capacités sont très sensibles aux variations de température et ils sont capables de détecter un gradient de 0,014°C par millimètre ! Sur la membrane plasmique du spermatozoïde, un canal calcique activable par la progestérone appelé CatSper stimule la nage flagellaire. CatSper est un complexe de 9 protéines : 4 qui forment un canal et 5 protéines régulatrices. Sachant que les cellules de la corona radiata entourant l’ovocyte sécrètent de la progestérone, cela permet d’expliquer la chémoattraction des spermatozoïdes.
Les cellules entourant l’ovocyte sécrètent également la protéine CRISP1 qui se lie aussi à CatSper (Ernesto et al., 2015). Il modulerait la nage des spermatozoïdes à proximité du l’ovocyte et des cellules environnantes pour la rendre plus efficace. Seuls 200 spermatozoïdes environ finiront par atteindre l’ovocyte. Par ailleurs, les spermatozoïdes peuvent survivre jusqu’à 6 jours dans les voies génitales femelles (Wilcox et al., 1995).
Contact et fusion entre les gamètes
La grande majorité des données classiques sur la fécondation des Mammifères a été obtenue in vitro. Or in vivo, des souris knock-out pour des acteurs jugés essentiels pour la fécondation in vitro se sont révélées être fertiles, ce qui a amené à corriger les modèles classiques.
Chez les Mammifères, la zone pellucide est une matrice glycoprotéique qui entoure les ovocytes et a une épaisseur moyenne de 17 µm. Elle est essentielle pour la fécondation (plus précisément pour la reconnaissance des gamètes et pour la prévention de la polyspermie), et pour la protection des embryons précoces avant l’implantation. La zone pellucide est composée essentiellement de glycoprotéines appelées ZP1 à ZP4. Chaque protéine ZP est un polypeptide qui est glycosylé de manière hétérogène avec des oligosaccharides liés à l’asparagine (N-) et à la sérine/thréonine (O-) qui, dans certains cas, sont sialylés et sulfatés. Chez certains mammifères, tels que le chat, la vache, le chien et le porc, ZP1 n’est pas présent et sa fonction est complètement remplacée par ZP4. Chez la souris, ZP4 n’est pas présent puisqu’il est codé par un pseudogène qui n’est pas exprimé lors de l’ovogenèse (Goudet et al., 2008). ZP1 et ZP4 proviennent de la duplication récente d’un gène ancestral et cela est suivi des vicissitudes habituelles dans ce cas avec de possibles transformations en pseudogènes ou délétion par redondance fonctionnelle (Smith et al., 2005). ZP2 est assez apparenté à ZP1 et ZP4. Par contre, ZP3 est évolutivement assez éloigné des 3 autres (Goudet et al., 2008).
Chez la souris, le knock-out ciblé soit du gène codant ZP2 soit du gène codant ZP3 empêche le développement d’une zone pellucide, conduisant à la stérilité. Des cas de stérilité dans une famille ont été reliés à une mutation dans ZP1 qui aboutit à une protéine aberrante qui reste dans le cytoplasme. Bien que normale, la protéine ZP3 qui s’associe à ZP1 reste également dans le cytoplasme et n’est pas secrétée (Huang et al., 2014), mettant en évidence la dépendance de ZP3 vis-à-vis de ZP1 pour sa localisation finale.
ZP2 et ZP3 semblent être les glycoprotéines sur lesquelles s’attache le spermatozoïde lors de son arrivée dans la zone pellucide. La comparaison des séquences de ZP3 de différentes espèces de mammifères révèle un degré élevé de divergence dans son domaine d’interaction avec le spermatozoïde par rapport à d’autres régions de la protéine. Cette divergence est attribuée à une sélection darwinienne positive et assure sans doute une importante barrière d’espèce (Williams et al., 2005). Lorsqu’on essaie de féconder des ovocytes de souris avec des spermatozoïdes humains, cela échoue mais lorsque ces ovocytes de souris expriment ZP2 humain alors cela réussit, montrant aussi l’importance du ZP2 dans les reconnaissances spécifiques des espèces (Baibakov et al., 2012).
La réaction acrosomiale est une exocytose dépendant du Ca2+ qui permet au spermatozoïde d’excréter le contenu de son acrosome. Dans les modèles classiques, elle était dépendante de l’interaction avec la zone pellucide. Il semble cependant que la réaction acrosomiale puisse avoir lieu avant, indépendamment de la zone pellucide.
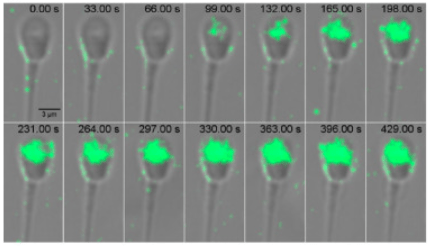
Source : https://www.jbc.org/article/S0021-9258(20)44442-4/fulltext
Cette réaction libère des enzymes notamment la hyaluronidase qui permet au spermatozoïde de se frayer un chemin à travers la corona radiata et la zone pellucide. Une autre enzyme libérée par la réaction acrosomiale, l’acrosine, joue un rôle plus controversé. Son inhibition par des anticorps chez le lapin et sa mutation perte-de-fonction chez le hamster par Crispr-Cas9 diminuent l’efficacité de la fécondation par les spermatozoïdes par ralentissement de la traversée de la zone pellucide mais des spermatozoïdes de souris ou de rats déficients en acrosine la traversent correctement (Hirose et al., 2020). Elle joue sans doute un rôle qui est compensable par une autre enzyme chez certaines espèces.
La réaction acrosomiale permet aussi de placer à la surface du spermatozoïde la protéine IZUMO qui sera importante pour la fusion avec l’ovocyte.
Des réactions acrosomiales trop précoces peuvent aboutir à de l’infertilité (Tesarik et Mendoza, 1995). Son déclenchement est donc étroitement régulé. Un pic de concentration d’AMPcyclique active la PKA (protéine kinase A) qui déclenche une cascade de signalisation aboutissant à l’activation de petites GTPases Rap1 et Rab3A qui remodèlent le cytosquelette et le trafic membranaire ce qui aboutit à la réaction acrosomiale (Pelletan et al., 2015).
La fusion des membranes plasmiques des 2 gamètes est initiée par l’interaction de IZUMO sur la membrane plasmique du spermatozoïde et de JUNO sur celle de l’ovocyte (Bianchi et al., 2014; Aydin et al., 2016).
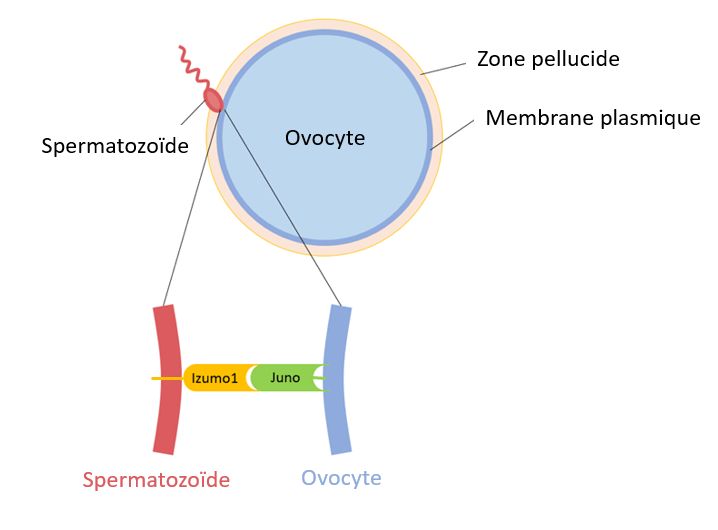
Les souris mâles déficientes en Izumo (mais pas les souris femelles) sont stériles car le sperme dépourvu d’Izumo ne peut pas fusionner avec les ovules. La zone pellucide des ovocytes Juno−/− est pénétrée in vivo par des spermatozoïdes de type sauvage, mais ceux-ci ne fusionnent pas avec l’ovocyte muté. Si on revient à la situation normale, après la fusion du spermatozoïde avec l’ovocyte, Juno est éliminé de la membrane plasmique ce qui constitue un mécanisme de blocage de la polyspermie (Bianchi et al., 2014).
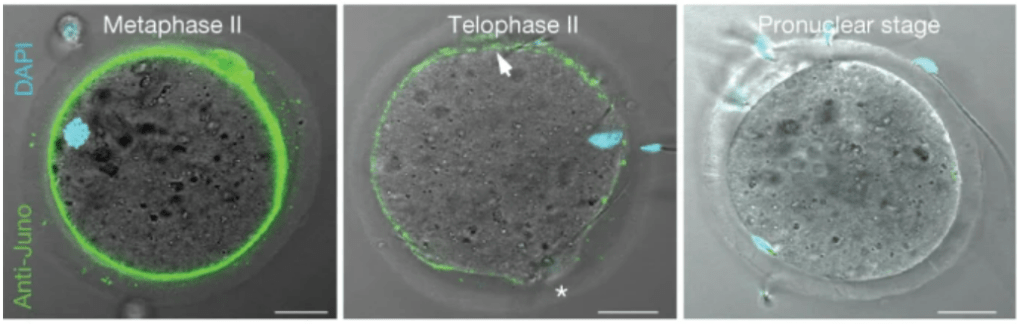
Source : https://www.nature.com/articles/nature13203
Depuis récemment, on sait que des mitochondries de la pièce intermédiaire du spermatozoïde passent dans le cytoplasme de l’ovocyte mais elles sont éliminées par autophagie. La transmission uniquement maternelle des mitochondries est ainsi préservée. Les mitochondries ont leur propre génome (avec 13 gènes codant des protéines mitochondriales et 24 gènes codant des ARN non codant chez l’humain) et les mutations de ce génome qui peuvent causer diverses maladies métaboliques (voir Russell et al., 2020) sont uniquement transmises par la mère (héritage maternel cytoplasmique).
Conséquences de la fécondation
Chez les amphibiens, l’arrivée du spermatozoïde se fait toujours par l’hémisphère animal. Elle entraîne un blocage de la polyspermie rapide sous la forme d’une dépolarisation membranaire. Une augmentation de la concentration du Ca2+ cytoplasmique provoque l’exocytose des granules corticaux.

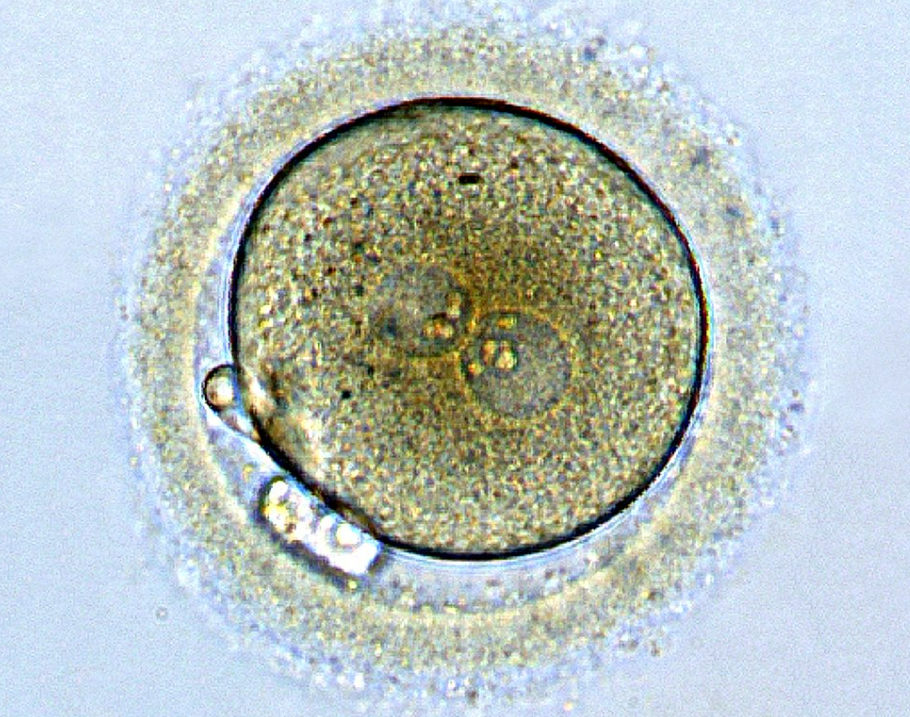
Les mucopolysaccharides des granules corticaux provoquent un appel d’eau dont l’afflux génère un espace entre la membrane plasmique et l’enveloppe vitelline, l’espace périvitellin : le zygote n’est plus relié à la gangue et à cause de la densité importante en vitellus du côté végétatif, il se tourne le pôle végétatif vers le bas et le pôle animal vers le haut (alors qu’auparavant son orientation était aléatoire). C’est la rotation d’équilibration qui a lieu habituellement 30 min après la fécondation. Lors de fécondations in vitro dans les laboratoires, c’est un bon indicateur que la fécondation a bien marché.
Les granules corticaux contiennent aussi des enzymes qui modifient la composition de la membrane vitelline et la transforme en membrane de fécondation (ou chorion). Le blocage de la polyspermie est donc d’abord ionique (dépolarisation), physique (espace périvitellin) et biochimique (composition de la membrane de fécondation).
Il existe une deuxième rotation qui a lieu 1h30 environ après la fécondation : la rotation de la couche superficielle du cytoplasme d’un angle de 30° environ vers le point d’entrée du spermatozoïde. C’est la rotation corticale. Les granules de mélanine sont entraînés par le cytoplasme mais certains restent en place formant un croissant gris. Ce croissant gris marque la future région dorsale de l’embryon.

Source : https://www.pnas.org/doi/10.1073/pnas.94.4.1224
Si on irradie la région végétative de l’ovocyte avec des UV, on perturbe la réaction corticale et on obtient des embryons tronqués, sans région dorsale et sans tête. Ceci est expliqué par le fait que lors de la rotation corticale, certaines molécules (protéines et ARNm) importantes pour la formation de la région dorsale sont entraînées du pôle végétatif vers le côté du croissant gris. Un deuxième axe (dorso-ventral) est ainsi formé, perpendiculaire à l’axe pôle animal-pôle végétatif qui avait été mis en place pendant l’ovogenèse. L’axe dorso-ventral n’est pas clairement séparé de l’axe antéro-postérieur de l’animal à ces stades précoces.
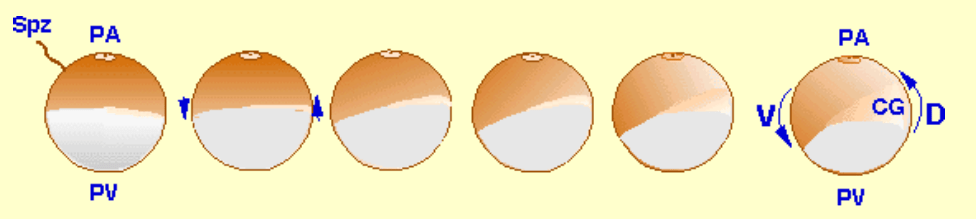
La rotation corticale dépend du réseau de microtubules. Après la fécondation, sous le contrôle du centriole amené par le spermatozoïde, des faisceaux de microtubules s’assemblent sous le cytoplasme cortical. Dans l’hémisphère végétatif, ces faisceaux sont parallèles.
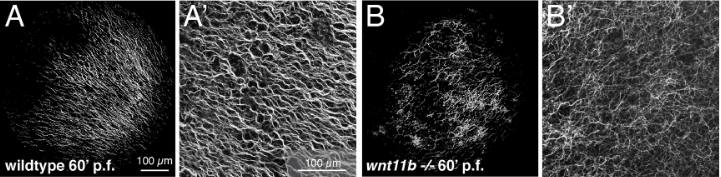
L’irradiation aux UV précédemment citée créé des liaisons covalentes entre le GTP et la tubuline et perturbe cette organisation. Un traitement à la colchicine ou au nocodazole perturbe aussi la formation des microtubules et inhibe la rotation corticale. On peut suivre les déplacements liés à la rotation corticale grâce à des microbilles fluorescentes.
Lors de la fécondation chez les Mammifères, l’entrée du spermatozoïde provoque une dépolarisation membranaire et une succession de pics de concentration en Ca2+ cytoplasmique (qui activent la protéine kinase dépendante de la calmoduline CaMKII (Knott et al., 2006)). Ces hausses de concentration calcique dépendent de PLC-ζ (PLC-zeta), une phospholipase C apportée par le spermatozoïde qui clive PI(4,5)P2 et permet de produire IP3 (inositol-triphosphate) qui active la sortie du Ca2+ du réticulum endoplasmique vers le cytosol (Wakai et al., 2011).
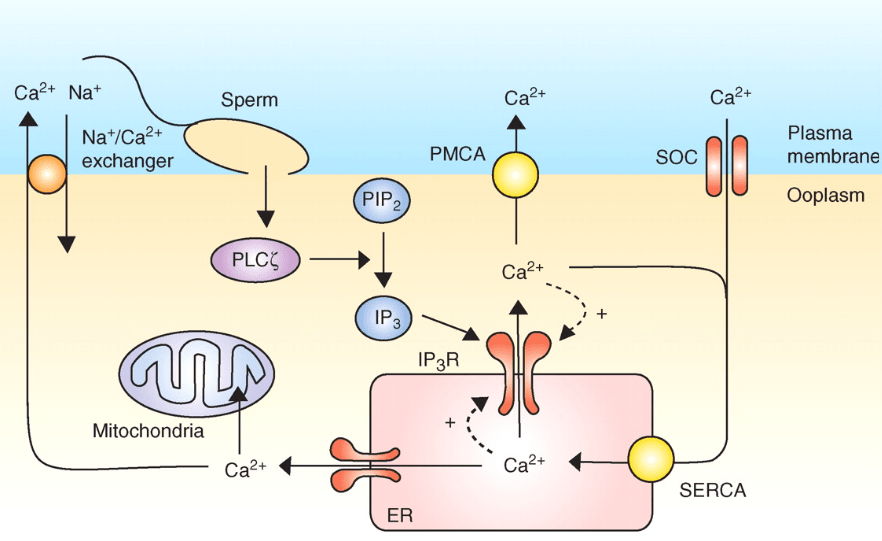
Source : https://cshperspectives.cshlp.org/content/3/4/a006767.long
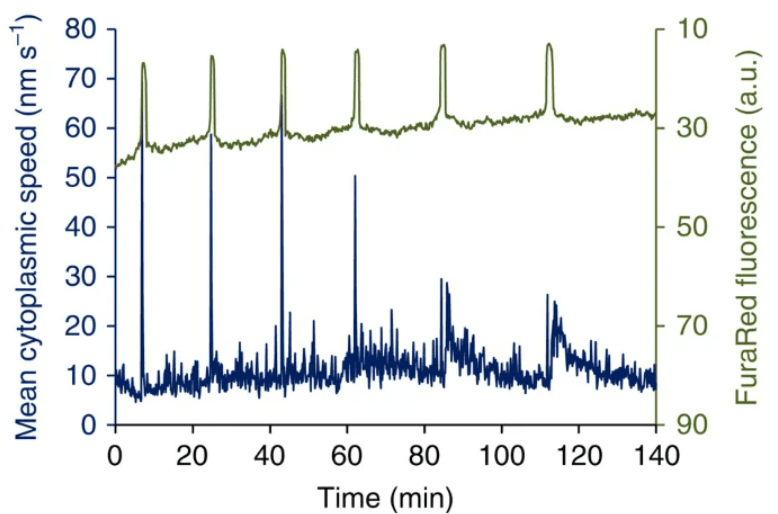
Une très forte concentration en Ca2+ cause la fermeture des canaux Ca2+ du réticulum endoplasmique et un repompage se fait d’où la forme en pic de ce signal. Plusieurs pics se succèdent. Ces pics calciques provoquent l’exocytose des granules corticaux et le déblocage de la méiose (dans ce dernier cas par stimulation de la dégradation du CSF (CytoStatic Factor). La méiose se termine avec l’expulsion du deuxième globule polaire.
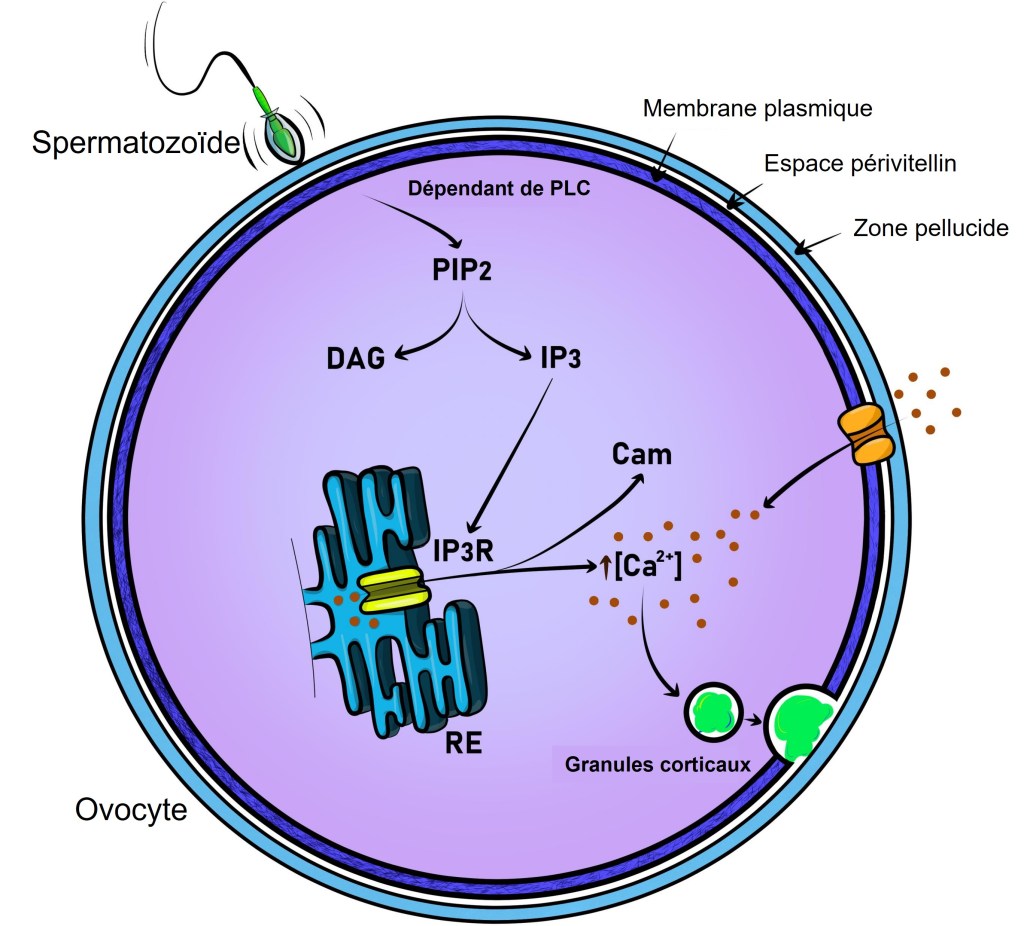
Seuls 4 pics calciques sont nécessaires pour déclencher l’exocytose des granules corticaux mais il en faut au moins 8 pour faire redémarrer la méiose (Ozil et al., 2005). Les enzymes dans les granules corticaux clivent les protéines ZP de la zone pellucide. Par exemple, la protéase ovastacine clive ZP2 qui perd sa capacité à interagir avec les spermatozoïdes. On observe aussi des mouvements cytoplasmiques rythmés qui dépendent de l’interaction actine-myosine et des pics de concentrations en Ca2+. Le blocage de ces mouvements cytoplasmiques aboutit à des développements anormaux mais leur fonction précise n’est pas encore claire (Ajduk et al., 2011).
Le matériel génétique paternel et maternel se trouvent dans des noyaux séparés appelés pronoyaux ou pronucléi (pluriel de pronucléus).
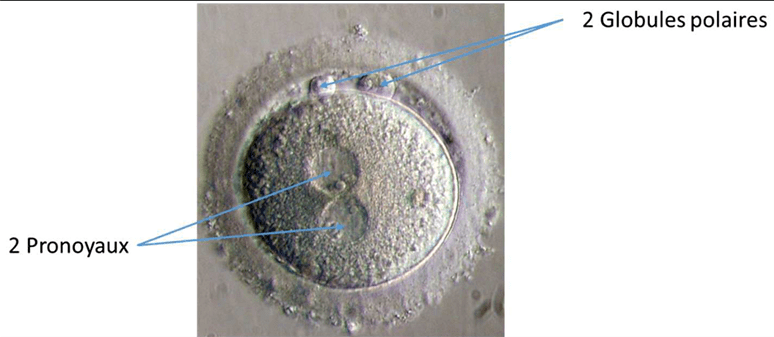
Dans le pronucléus mâle, les protamines sont remplacées par des histones maternelles sous la direction de la protéine Hira (Loppin et al., 2005). Chaque pronucléus fait réplication à part (8h après la fécondation chez la souris) et ce n’est que lors de la métaphase de la première mitose que les chromosomes maternels et paternels se retrouvent côte à côte. Ainsi, les premiers noyaux vraiment diploïdes sont ceux de l’embryon au stade 2 cellules.
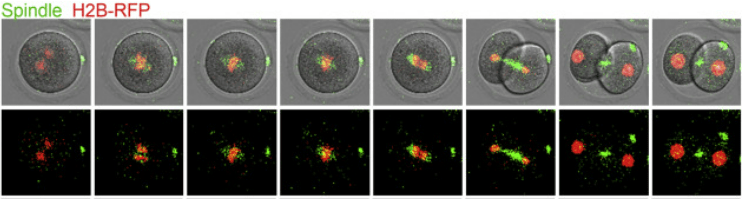
Les particularités de la fécondation chez la drosophile. La drosophile présente quelques différences notables avec les fécondations décrites ici. L’activation de l’ovocyte a lieu lors de l’ovulation, avant même que l’arrivée des spermatozoïdes. Des canaux calciques s’ouvrent et les concentration en Ca2+ augmentent dans le cytoplasme, indépendamment du spermatozoïde. Celui-ci ne peut féconder qu’une région bien précise de l’ovule, celle qui se trouve sous le micropyle qui est un tunnel creusé dans la paroi de l’œuf (chorion). Elle correspond à la future région dorsale antérieure. Le micropyle ne peut être traversé que par un spermatozoïde à la fois, ce qui limite les risques de polyspermie. Il n’y a pas d’exocytose de granules corticaux.
La procréation médicalement assistée (PMA)
L’Organisation Mondiale de la Santé (OMS) définit l’infertilité par l’absence de grossesse après au moins un an de rapports sexuels réguliers non protégés chez les couples en âge de procréer. Elle affecte entre 9% et 15% des couples selon les études (Boivin et al., 2007).
La PMA apporte une réponse à la stérilité ou à l’infertilité et représentait 3,4% des naissances en France en 2018 (contre 2,6% en 2009). Plusieurs millions de personnes sont nés dans le monde grâce à la PMA depuis la mise au point de la fécondation in vitro en 1978.
Selon le type d’infertilité, il y a plusieurs options de traitement : thérapie médicamenteuse (induction de l’ovulation), insémination intra-utérine, fécondation in vitro (FIV) et transfert embryonnaire (FIVETE), injection intracytoplasmique de spermatozoïde (ICSI).
L’insémination intra-utérine est la plus pratiquée dans le monde. Elle consiste à déposer une préparation enrichie en spermatozoïdes mobiles directement dans la cavité utérine et elle est prescrite en cas d’infertilité masculine avec des spermatozoïdes avec un taux de mobilité normal ou subnormal.
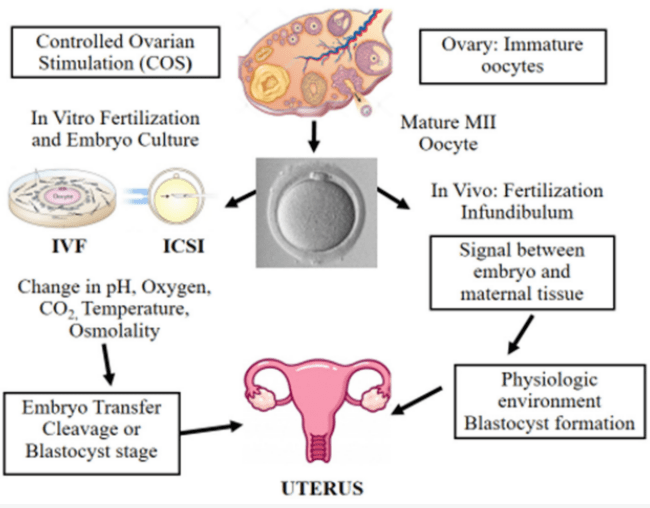
La première naissance d’un enfant issu du FIV a eu lieu en 1978 (Louise Brown en Angleterre). En France, cela est arrivé avec Amandine en 1982. La procédure est la suivante : une stimulation hormonale ovarienne et un suivi du développement des follicules par échographie. Les ovocytes sont prélevés quelques heures avant l’ovulation puis mis en présence des spermatozoïdes préparés. Les embryons obtenus sont cultivés in vitro pendant 2 à 6 jours afin d’évaluer leur bon développement par leur morphologie. Des embryons ainsi sélectionnés sont transférés dans l’utérus maternel (le taux de succès de l’implantation et d’une grossesse par embryon est de 20 à 30% (de Mouzon et al., 2012). Les embryons surnuméraires et qui ont eu une bonne morphologie peuvent être congelés pour stockage.
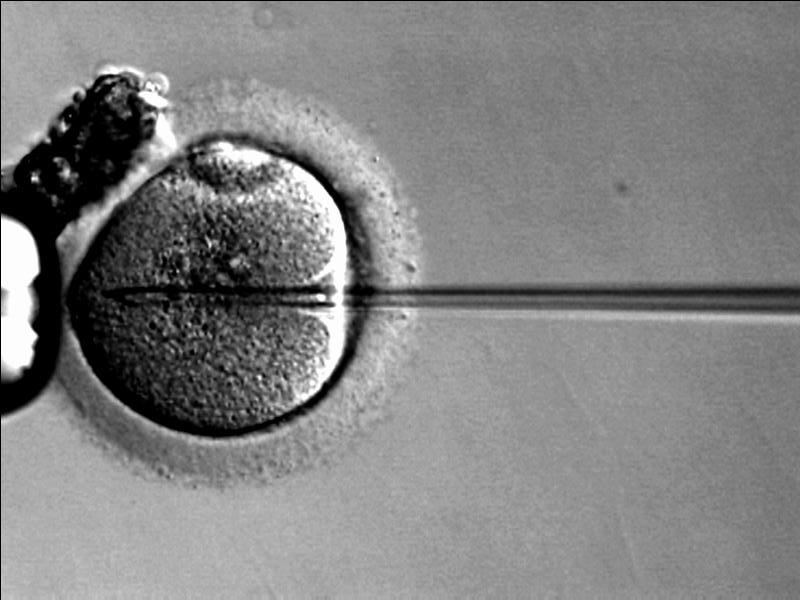
Pour l’ICSI, on charge dans une micropipette un spermatozoïde dont on casse le flagelle et on l’injecte dans le cytoplasme ovocytaire. La première ICSI a été réalisée en Belgique en 1992. Les indications de cette technique sont de graves défauts de mobilité des spermatozoïdes, des anomalies de la zone pellucide, des anomalies des capacités fusiogènes des membranes plasmiques des gamètes et des anomalies de l’exocytose des granules corticaux (pouvant mener à la polyspermie). La technique est devenue très utilisée, jusqu’à 2/3 des procédures de PMA dans certains pays (Dyer et al., 2016).
Même si toutes ces procédures sont souvent couronnées de succès, elles peuvent provoquer des modifications dont l’impact au cours de la vie des bébés nés grâce à la PMA est encore à évaluer : par exemple, les télomères des bébés conçus par PMA sont un peu plus courts que les enfants conçus naturellement (Wang et al., 2022).
> Après la fécondation, a lieu le clivage.
LA CARTE MENTALE :

LES MOTS CROISES POUR REVISER :
EN DIRECT DES LABOS :
AUTRES RESSOURCES SUR LE SUJET :
Sur la gamétogenèse et la fécondation
- Adhérences cellule-cellule
- Arabidopsis thaliana
- Axe antéro-postérieur chez la drosophile
- Biomécanique du développement
- Caenorhabditis elegans
- Concepts principaux
- Contrôle de la traduction
- Contrôle de la transcription
- Contrôle génétique et épigénétique
- Croissance du tube pollinique et double fécondation chez les Angiospermes
- Croissance et guidage axonal
- Des modèles animaux moins classiques
- Développement de l’oeil des Vertébrés
- Développement et évolution
- Et l’Humain ?
- Exercices sur l’ovogenèse, la spermatogenèse et la fécondation
- Exercices sur le contrôle de l’expression des gènes
- Exercices sur le développement des bourgeons de membre
- Exercices sur le développement des muscles striés squelettiques
- Exercices sur le développement des végétaux et les hormones végétales
- Exercices sur les cycles et les divisions cellulaires
- Exercices sur les étapes du développement, les inductions embryonnaires et la mise en place des axes de polarité
- Exercices sur les matrices extracellulaires, le cytosquelette et les adhérences cellule-cellule
- Exercices sur les voies de signalisation
- Glossaire
- Glossaire des termes liés à la génétique
- Glossaire des termes liés au cytosquelette, la matrice extracellulaire, l’adhérence et la migration cellulaire
- Hématopoïèse et développement des cellules du système immunitaire
- Histoire de la biologie cellulaire et de la biologie du développement
- L’acide rétinoïque
- L’apoptose
- L’autophagie
- L’organogenèse
- L’ovogénèse prépare le développement embryonnaire
- La drosophile
- La formation des somites
- La gastrulation
- La gastrulation (version allégée)
- La métamorphose chez les Hexapodes et les Amphibiens
- La neurogénèse chez les mammifères adultes
- La neurulation
- La poule
- La signalisation calcique
- La souris
- La superfamille TGFβ et ses voies de signalisation
- La voie de signalisation de l’auxine et ses rôles
- La voie de signalisation Hedgehog
- La voie de signalisation Hippo et ses composants YAP/TAZ
- La voie de signalisation Notch
- Le clivage
- Le cytosquelette
- Le destin des cellules et les réseaux de régulation génique
- Le développement des bourgeons de membre
- Le développement des muscles striés squelettiques
- Le développement des organes génitaux et des cellules germinales
- Le développement du cortex
- Le méristème apical caulinaire en phase végétative et lors de la formation d’une fleur
- Le poisson zèbre
- Le xénope
- Les cellules des crêtes neurales
- Les cellules et les gènes en action dans le développement
- Les cellules souches
- Les cellules tumorales
- Les cycles et les divisions cellulaires
- Les étapes du développement
- Les étapes du développement embryonnaire d’Arabidopsis thaliana et leur contrôle
- Les inductions embryonnaires et les gradients de morphogène
- Les matrices extracellulaires animales
- Les organismes modèles
- Les outils pour étudier l’expression et la fonction des gènes
- Les parois des cellules végétales
- Les techniques et les outils pour la biologie cellulaire
- Les transitions épithélio-mésenchymateuses et les migrations cellulaires
- Les vésicules extracellulaires
- Les voies de signalisation
- Les voies de signalisation FGF
- Mise en place des axes chez les Vertébrés
- Structures et processus cellulaires
- Voies de signalisation WNT
