
Par Patrick Pla, Université Paris-Saclay
- Introduction
- Le développement des neurones corticaux
- Le développement des cellules gliales
- Régionalisation au sein du cortex
- Mise en place, maintien et modifications des connexions
- Produire in vitro des neurones corticaux à partir de cellules souches
Introduction
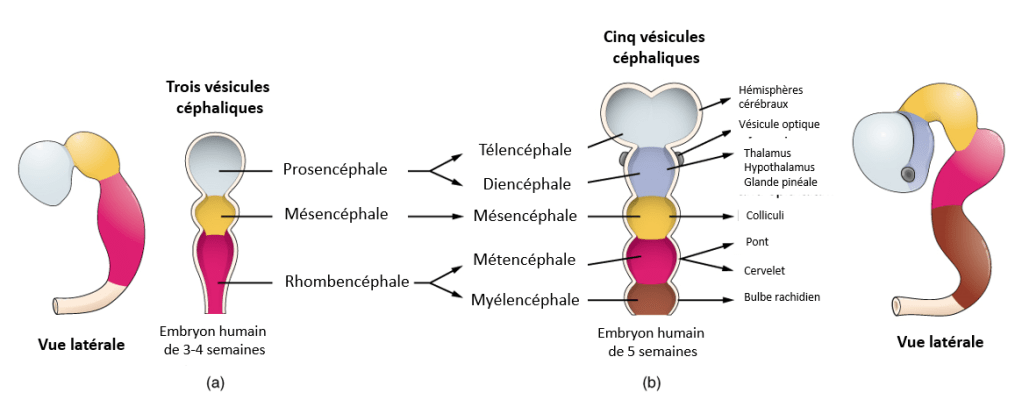
Le néocortex cérébral (qu’on abrège souvent en cortex) se développe dans la zone la plus superficielle dorsale du télencéphale des mammifères. Le télencéphale est l’une des 5 vésicules céphaliques des Vertébrés et c’est la plus antérieure. Il se scinde en deux hémisphères qui possèdent chacune une cavité : le ventricule I et le ventricule II.
Le développement des neurones corticaux
Généralités
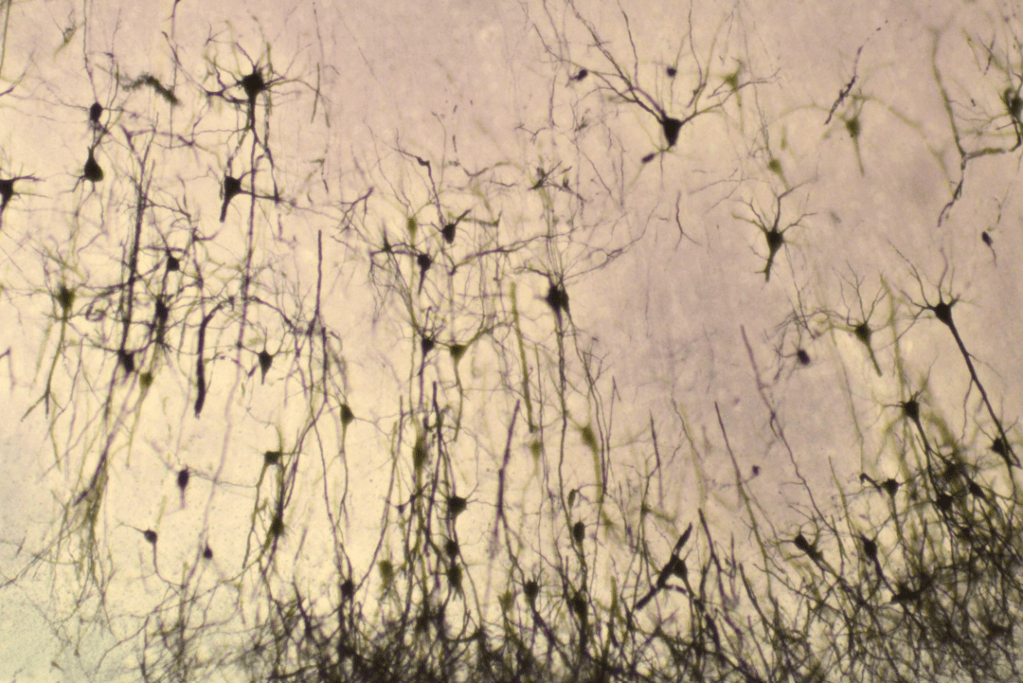
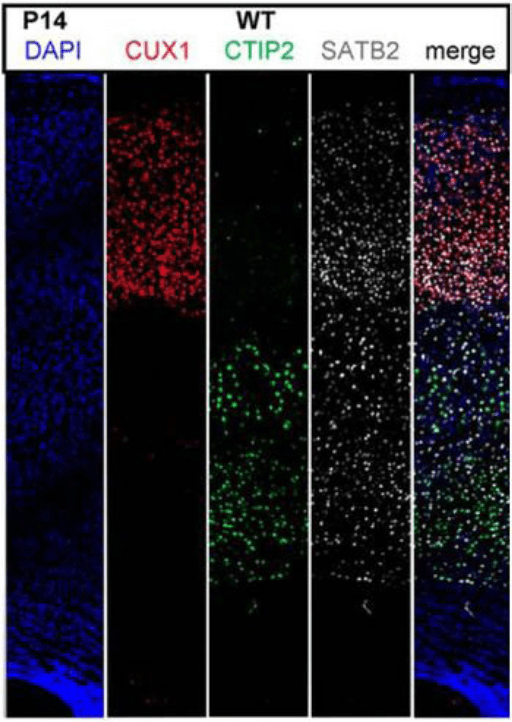
Le néocortex est constitué de deux classes principales de neurones : les neurones glutamatergiques (excitateurs) et les interneurones GABAergiques (inhibiteurs). Les neurones glutamatergiques excitateurs représentent 70 à 80 % des neurones qui peuplent le néocortex humain. La plupart d’entre eux sont des neurones pyramidaux, caractérisés par leur morphologie : un corps cellulaire conique, des arborisations dendritiques apicales et basales qui portent des épines dendritiques comme principales structures postsynaptiques excitatrices, et un axone se projetant généralement vers des cibles à longue distance. Les neurones pyramidaux se répartissent en 6 couches et sur chaque couche, ils présentent des patrons uniques d’expression génique, de morphologie et de connexions . Schématiquement, les neurones pyramidaux plus profonds (couches corticales V et VI) se projettent principalement vers les structures sous-corticales, et les neurones pyramidaux supérieurs (couches corticales II et III) se projettent principalement vers d’autres zones corticales (projections cortico-corticales), tandis que les neurones de la couche IV constituent la principale entrée d’informations en provenance des régions sous-corticales.
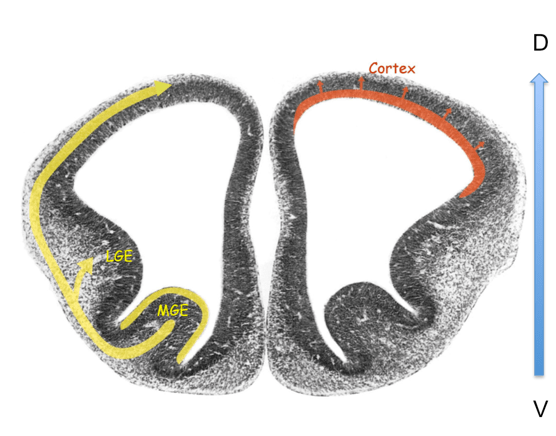
Les interneurones inhibiteurs GABAergiques, qui n’envoient généralement que des projections axonales locales, constituent environ 25 % des neurones du cortex humain et sont subdivisés en de nombreux types selon l’expression des gènes, la morphologie, la connectivité et les propriétés physiologiques. Ces interneurones ne sont pas générés sur place, contrairement aux neurones excitateurs. Ils sont générés ventralement à partir des éminences ganglionnaires médiales et leurs précurseurs migrent ensuite dans le cortex (Chung et al., 2025).
Tout comme les neurones excitateurs, les neurones inhibiteurs du cortex présentent une grande diversité. Grâce à des études transcriptomiques en cellules uniques (scRNAseq) on commence à bien connaître l’ampleur de cette diversité (Gouwens et al., 2020).
Enfin, le néocortex est morcelé en de nombreuses aires corticales, chacune spécialisée dans des fonctions spécifiques, liées à leur connectivité entre elles et avec le reste du cerveau. Les neurones de différentes zones corticales présentent également des patrons distincts d’expression génique et de morphologie, ainsi que des proportions spécifiques de classes neuronales.
Les cellules souches neurales du cortex
Les neurones (puis les cellules gliales, nous le verrons plus loin) sont générés à partir de cellules souches neuronales qui sont en fait des cellules de la glie radiaire qualifiée de apicale (aRG) car leur prolongement est en contact avec la partie apicale du neuroépithélium (du côté du ventricule, c’est-à-dire de la lumière du tube neural). La polarité de ces cellules neuroépithéliales est très importante, ce qui est illustré par le fait que des perte-de-fonctions de gènes codant des protéines impliquées dans la structure des jonctions serrées et des jonctions adhérentes qui sont essentielles pour le maintien de la polarité aboutissent à des développements anormaux du cortex tels que des microcéphalies ou favorise la survenue de l’autisme et de crises d’épilepsie (O’Driscoll et al., 2010, Bendriem et al., 2019; Sterling et al., 2020).
Ces cellules souches peuvent subir une division symétrique qui génère deux cellules souches (expansion) ou une division asymétrique où l’une des cellules reste cellule souche (auto-renouvellement) et l’autre devient un précurseur qui peut proliférer mais qui finira par se différencier (en neurone ou en cellule gliale). La régulation entre division symétrique ou asymétrique dépend de nombreux paramètres, notamment de l’orientation du fuseau mitotique et, étonnamment, de la taille des mitochondries (les cellules qui gardent des caractères de cellule souche ont de longues mitochondries tubulaires, les cellules qui perdent ces caractères ont des mitochondries plus nombreuses mais plus petites) (Iwata et al., 2021).
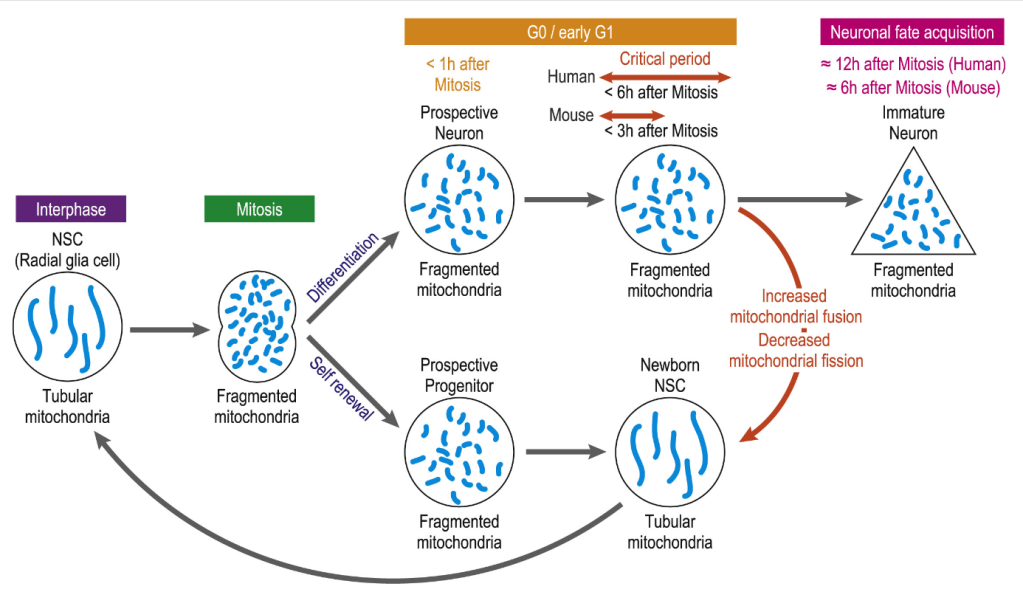
Un dysfonctionnement de la division des cellules de la glie radiaire peut mener à la microcéphalie.
Les cellules de la glie radiaire ont un cil primaire qui baigne dans le liquide céphalo-rachidien. Des études de protéomique ont montré que ce liquide a une composition qui varie au cours du temps : il est d’abord enrichi en Sonic Hedgehog (à partir de E10,5 chez la souris) puis sa concentration diminue tandis que la concentration en acide rétinoïque augmente (vers E14,5) (Chau et al., 2015). L’auto-renouvellement et la prolifération des cellules souches neurales sont dépendants de molécules présentes dans le liquide céphalo-rachidien.
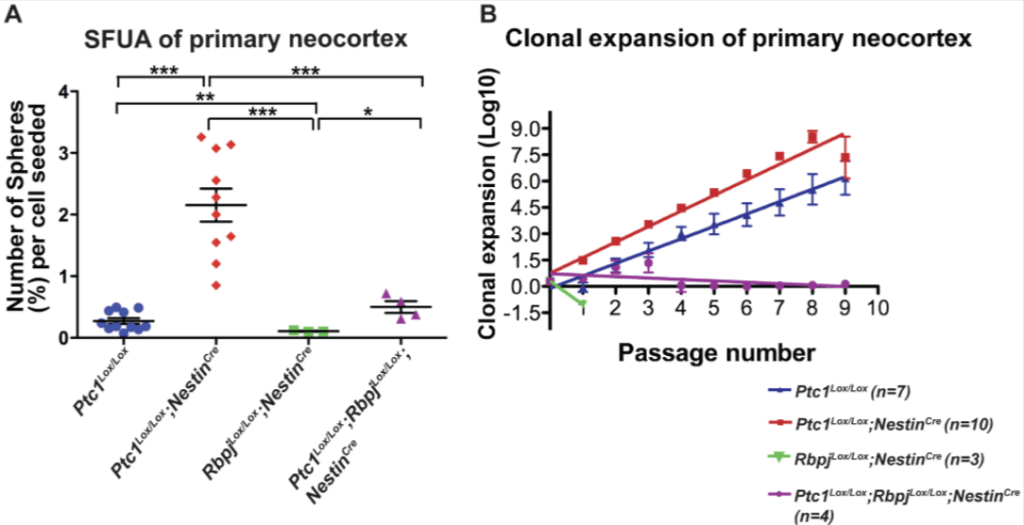
(A) Les souris Ptc1Lox/Lox;NestinCre (n = 10) présentent une activité Sonic Hedgehog élevée et constitutive dans les cellules souches neurales car on y a délété le gène codant Patched1 qui est un inhibiteur de cette voie. On génère des neurosphères à partir du cortex de ces souris (et de témoins ainsi que de souris RbpjLox/Lox;NestinCre qui ne peuvent plus activer la voie Notch dans les cellules souches neurales). La suractivation de la voie Shh aboutit à une augmentation de 8 fois du nombre de colonies de neurosphères primaires par cellule ensemencée par rapport aux témoins de la même portée. Cependant la voie Notch doit concomitamment être fonctionnelle. (B) Expansion clonale des neurosphères dérivées de cortex à E14.5. Le nombre de cellules obtenues à chaque passage a été exprimé en logarithme (Log10) et une analyse de régression linéaire a été réalisée sur les courbes obtenues. Le taux d’expansion clonale des neurosphères est représenté par la pente de la ligne. Le nombre de cellules souches est significativement plus élevé dans les neurosphères Ptc1Lox/Lox;NestinCre par rapport aux neurosphères Ptc1Lox/Lox. Cependant, une fois de plus, la voie Notch doit pouvoir être activée. Source : https://journals.plos.org/plosone/article?id=10.1371/journal.pone.0014680
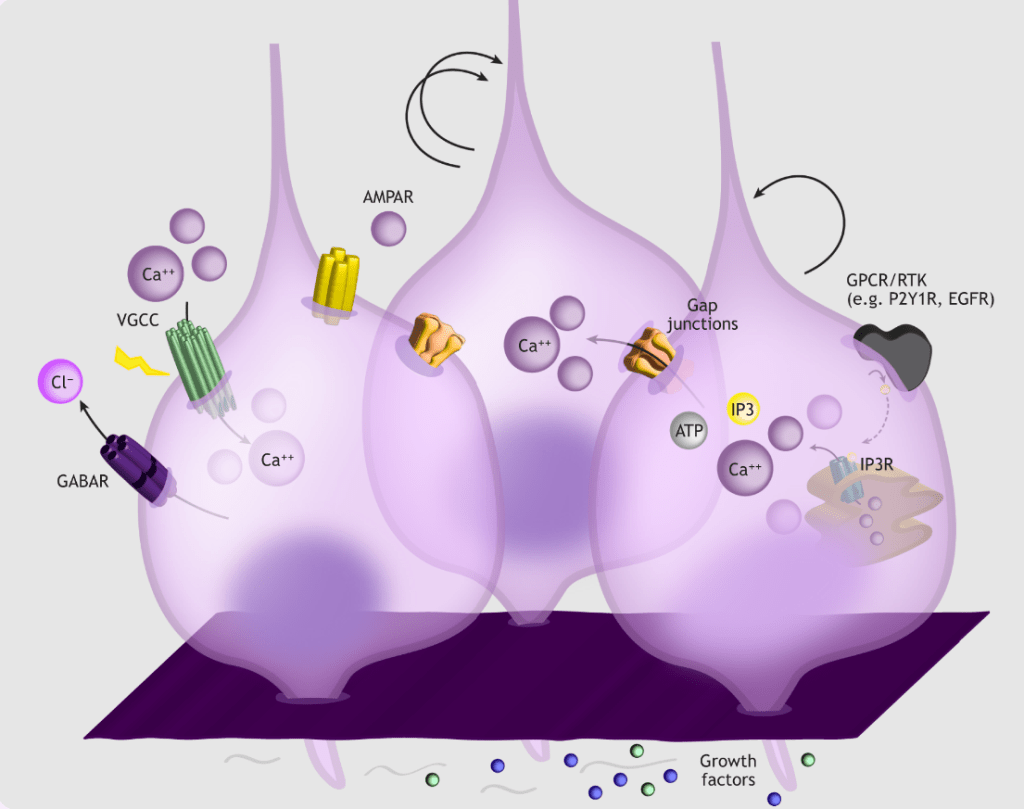
La prolifération des cellules de la glie radiaire est aussi sous le contrôle de la signalisation calcique (McKinney et al., 2022).
Des cellules souches neurales du cortex aux neurones différenciés
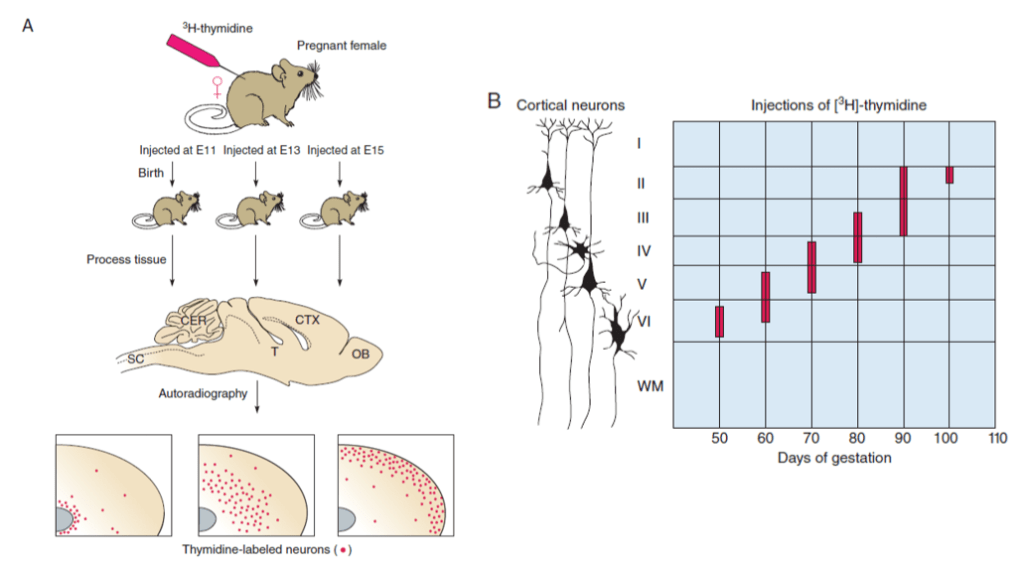
Le néocortex des Mammifères possède 6 couches avec des types neuronaux bien précis dans chacune de ces couches.
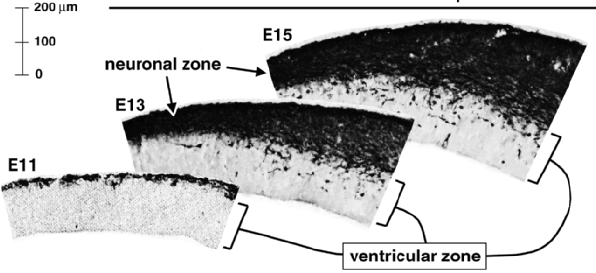
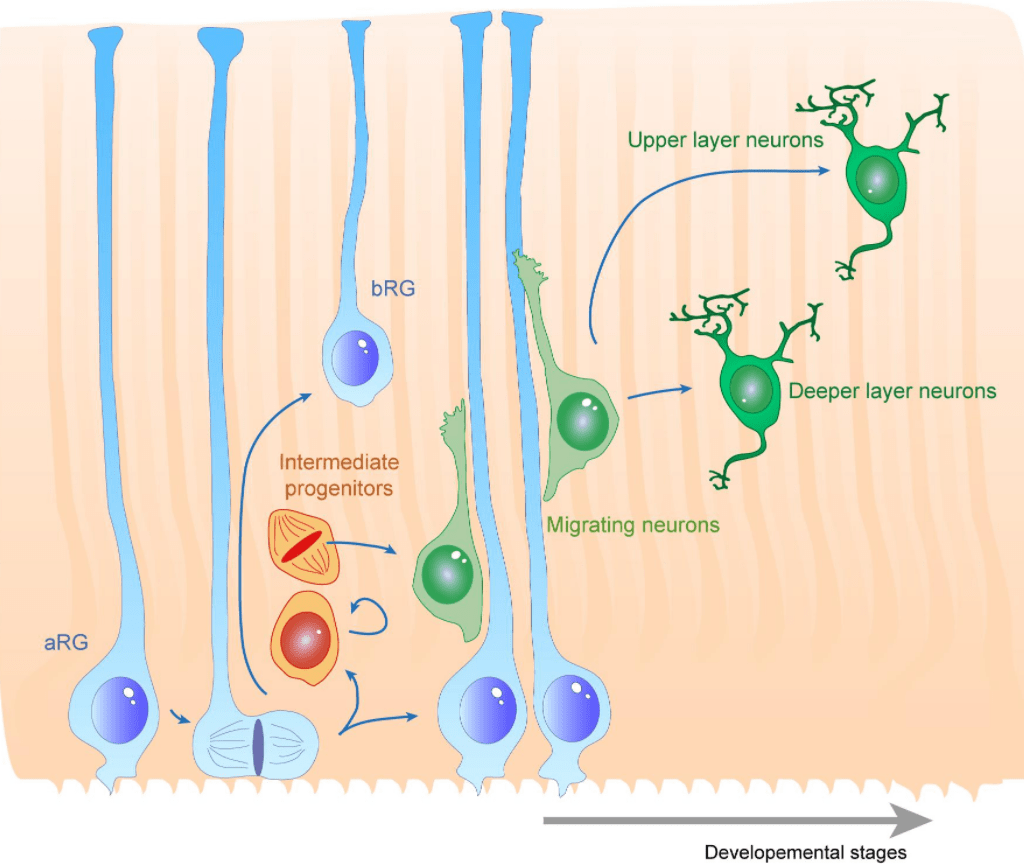
Les 6 couches sont générées de manière inversée. Les neurones les plus précoces occupent les couches les plus profondes et les derniers neurones produits occupent les couches les plus superficielles. Les neurones se développent par vagues de neurogenèse, migration radiale et différenciation à partir des progéniteurs gliaux radiaux de la zone ventriculaire et des cellules progénitrices intermédiaires de la zone sous-ventriculaire.
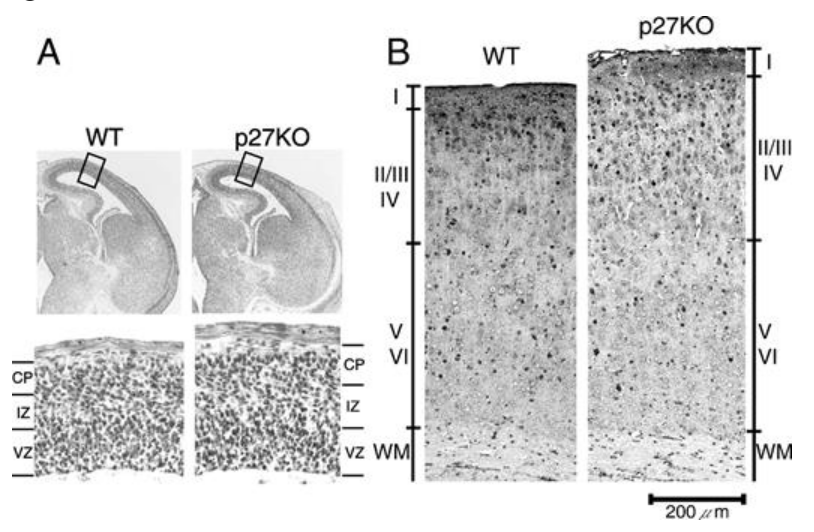
La vitesse de prolifération des cellules souches et des précurseurs tend à ralentir au cours du temps durant le développement du cortex. Si on abolit l’expression du gène p27 qui est un inhibiteur de CDK qui « freine » la progression dans le cycle cellulaire, on obtient des couches corticales II-IV qui possèdent plus de neurones et qui sont plus épaisses (Caviness et al., 2003).
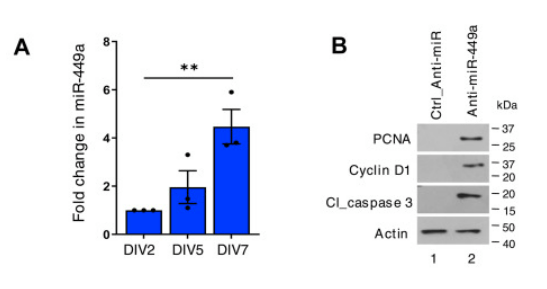
Des régulations traductionnelles ont lieu comme le montre le rôle joué par le microARN miR-449a dans l’arrêt de la prolifération des précurseurs neuronaux. miR-449a cible les ARNm de CDC25A et de Cycline D1 qui codent des protéines qui favorisent la prolifération (Chauhan et al., 2024).
Durant tout le processus, le maintien des cellules souches neurales est crucial. Les progéniteurs neurogéniques intermédiaires expriment Delta-like 1 (Dll1) et activent la voie Notch dans les cellules souches ce qui inhibe leur différenciation et les maintient dans un état de cellule souche (l’importance de cette signalisation a déjà été mise en évidence dans les expériences en parallèle avec la voie Shh plus haut). Cette communication Dll1/Notch juxtacrine dépend des longs prolongements avec lesquels les progéniteurs et les cellules souches entrent en contact (Nelson et al., 2013).
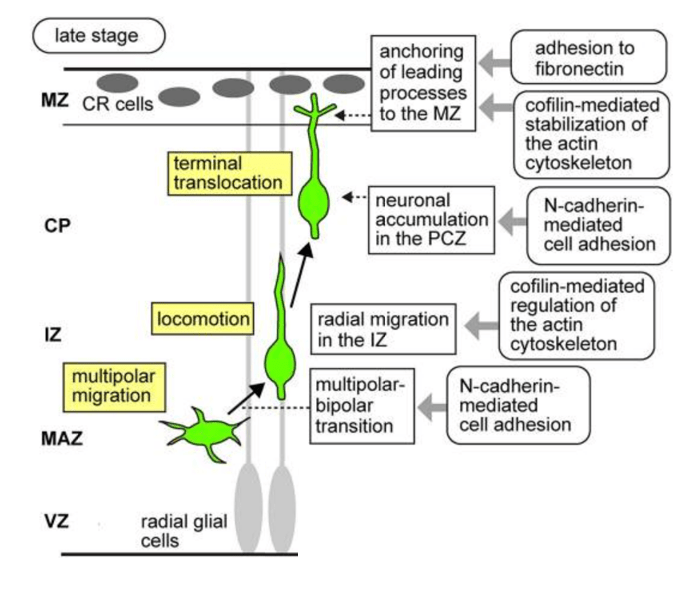
Les neurones migrent d’abord de manière multipolaire avant de prendre une forme bipolaire et de migrer le long de la glie radiaire. La transition entre ces deux phases dépend du récepteur Unc5D des neurones qui interagit avec le Glypican3 (ou GPC3), une protéine extracellulaire attachée à la membrane plasmique des cellules de la glie radiaire par une ancre GPI (Akkermans et al., 2022). Les patients avec des mutations dans le gène codant GPC3 présentent des anomalies du squelette mais aussi des malformations des couches corticales dûes à un retard de migration des futurs neurones.
La migration radiale des nouveaux neurones est réalisée par la succession de plusieurs cycles de formation d’une protrusion frontale suivie par une nucléokinèse, c’est-à-dire le déplacement du noyau à l’intérieur du cytoplasme. Tous ces mouvements dépendent d’une importante coordination entre la dynamique des microfilaments et des microtubules.
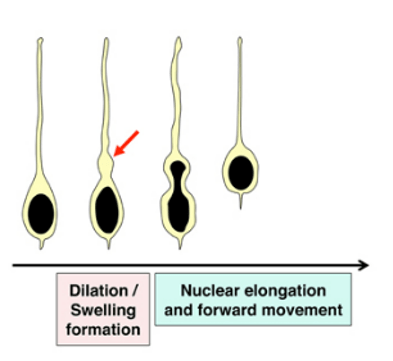
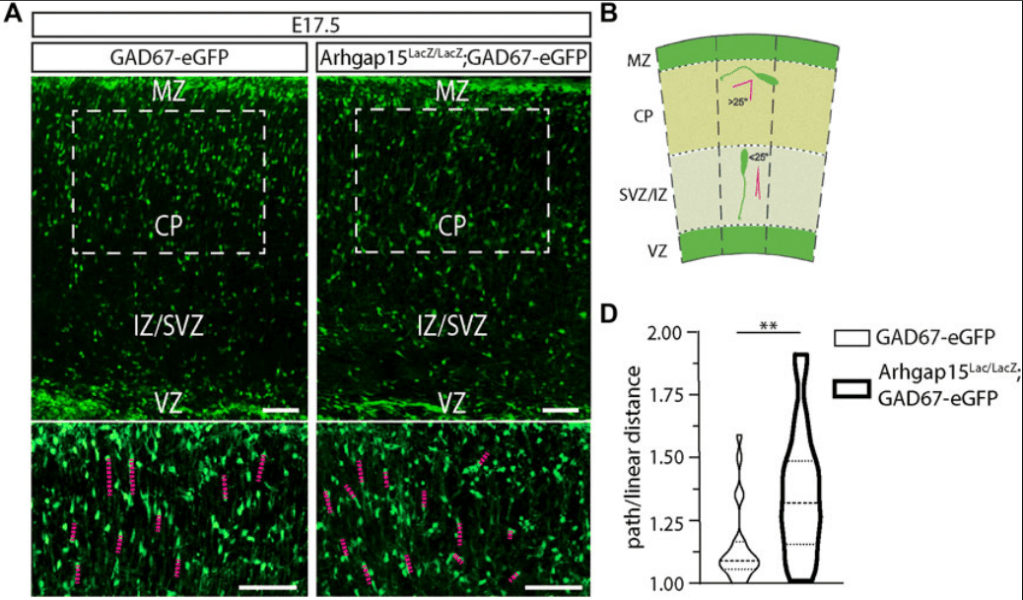
Des protéines contrôlant le cytosquelette tels que les petites GTPases Rho, Rac et Cdc42 sont importantes pour la migration radiale des nouveaux neurones (qu’ils soient excitateurs ou inhibiteurs). Par exemple, Rac1 est exprimé à l’avant de ces neurones et sa fonction est contrôlée par la protéine de type GAP (GTPase-Activating Protein) ARHGAP15 qui inhibe Rac1. En absence d’ARHGAP15, les futurs neurones ont une migration qui n’est pas seulement radiale mais multidirectionnelle ce qui les amène à des destinations anormales. ARHGAP15 agit en limitant la capacité de Rac1 à produire des prolongements cellulaires exploratoires, ce qui aboutit à une meilleure directionalité de la migration (Liaci et al., 2022).
Chez le mutant de souris reeler, les nouveaux neurones sont incapables de passer par-dessus les autres pour générer des couches plus externes du cortex comme c’est le cas habituellement. La mutation a été isolée dans le gène codant la Reeline, une large glycoprotéine de plus de 3.000 acides aminés. Elle est secrétée par les cellules les plus superficielles du cortex : les cellules de Cajal-Retzius. La Reeline est perçue par les nouveaux neurones grâce à deux co-récepteurs ApoER2 (ou LRP8) et VLDLR et elle contrôle leur migration (Feng et al., 2007).
Les nouveaux neurones en migration présentent des oscillations de concentration intracellulaire en Ca2+ qui sont nécessaires à cette migration. Ces oscillations sont activées en partie par, d’une part, la liaison du facteur neurotrophique BDNF sur son récepteur TrkB (via IP3) et d’autre part, par la liaison du glutamate sur les récepteurs NMDA (via l’ouverture de canaux calciques voltage-dépendants) (McKinney et al., 2022).
Chez l’Homme, des mutations dans de nombreux gènes affectant la migration neuronale aboutissant à des défauts plus ou moins prononcés de la morphologie et de l’anatomie du cortex ont été caractérisées.
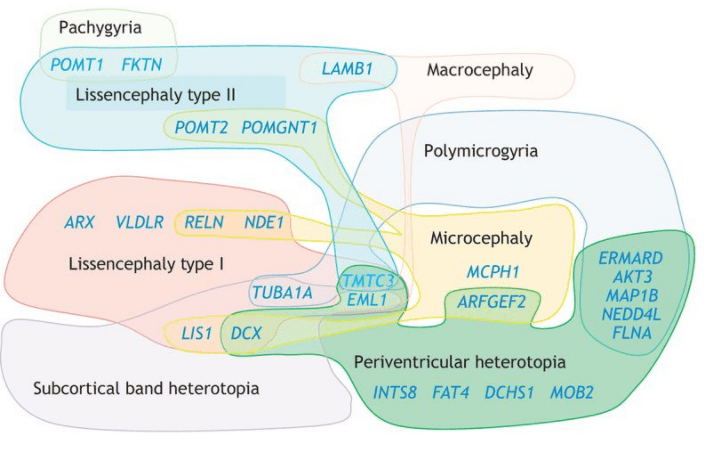
La migration des interneurones corticaux GABAergiques des éminences ganglionnaires médiales jusqu’au cortex en position plus dorsale (une migration qualifiée de tangentielle) a été moins étudiée que la migration radiale des neurones d’origine corticale. Néanmoins, des protéines importantes pour cette migration ont été caractérisées telles celles du complexe formant un canal calcique voltage-dépendant de type L (LTCC) (Bortone et Polleux, 2009). Des variants génétiques qui altèrent la fonction des LTCC ont été impliqués dans divers troubles neuropsychiatriques (Bhat et al., 2012). Par exemple, les mutations de gain de fonction dans le gène CACNA1C, qui code l’isoforme Cav1.2, provoquent le syndrome de Timothy, un trouble neurodéveloppemental rare associé à l’autisme et à l’épilepsie (Splawski et al., 2004). En utilisant des assembloïdes (plusieurs organoïdes accolés) de cerveau antérieur dérivés de patients atteints du syndrome de Timothy, des chercheurs ont mis en évidence un défaut de migration des interneurones corticaux GABAergiques (Birey et al., 2017).
Les progéniteurs qui deviennent post-mitotiques se différencient ensuite dans un sous-type de neurone qui dépend de la couche corticale occupée et se branchent menant à la mise en place de synapses fonctionnelles. Par exemple, FEZ Family Zinc Finger 2 (Fezf2) est un régulateur et sélecteur transcriptionnel requis pour la spécification des sous-types de neurones de projection corticospinale qui se trouvent dans la couche V (Molyneaux et al., 2005). Fezf2 active l’expression du récepteur de guidage axonal EphB1 et du transporteur vésiculaire du glutamate 1 (vGlut1) dans ces neurones. Fezf2 est également nécessaire pour leur développement dendritique. Pour ce faire, il active l’expression d’une batterie de microARNs. Le cluster miR-193b et miR-365 est exprimé de manière différentielle durant le développement des neurones de projection corticospinaux en comparaison avec les neurones calleux, et ce cluster réprime l’expression de MAPK8 qui est exprimé dans les neurones projetant vers le corps calleux (Lyer et al., 2021). Le gain de fonction des miARN miR-193b et miR-365 contrôle la ramification dendritique dans les neurones de projection corticale et phénocopie la perte de fonction de MAPK8. Dans la couche V, Fezf2 inhibe également l’expression de Tbr1 qui est nécessaire pour la détermination des neurones corticothalamiques (qui se développent dans la couche VI) et celle de Satb2 qui est nécessaire pour la détermination des neurones qui projettent vers le corps calleux (Chen et al., 2008). A l’inverse, Tbr1 et Sox5 sont tous deux exprimés à des niveaux élevés dans les neurones de projection corticothalamiques dans la couche VI. Ils favorisent ce destin neuronal en réprimant directement l’expression de Fezf2 dans cette couche (Han et al., 2011; McKenna et al., 2011).
Résumé du paragraphe précédent

Des études in vitro et in vivo ont montré que le gène Arx code un facteur de transcription généralement répresseur qui joue un rôle important pour le développement du cerveau (Seufert et al., 2005). Parmi les rôles de ce gène figurent la régionalisation du cerveau, la prolifération des progéniteurs corticaux, la migration des interneurones inhibiteurs à GABA et l’engagement des futurs neurones cholinergiques dans leur voie de différenciation (Colombo et al., 2004; Marsh et al., 2016; Friocourt et Parnavelas, 2010). De nombreuses mutations du gène ARX ont été signalées dans plus d’une douzaine de troubles neurologiques précoces différents, où des déficiences intellectuelles sont associées parfois à des crises d’épilepsie (notamment le syndrome XLAG qui comporte une lissencéphalie (=absence de circonvolutions)).
Aspects évo-dévo
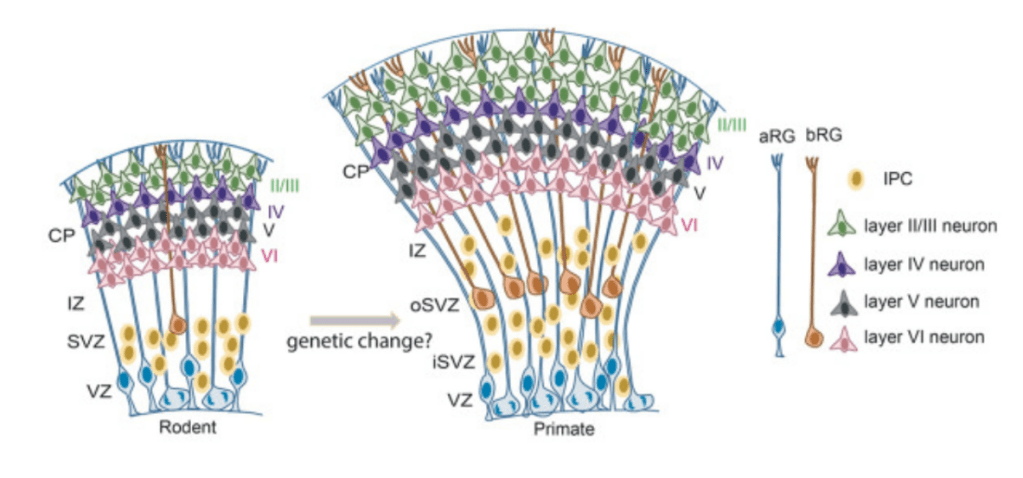
Avec plus de 80 milliards de neurones, l’humain est l’une des espèces de Mammifères possédant le plus grand nombre de neurones cérébraux (bien qu’il soit nettement inférieur en nombre à celui des éléphants et des baleines), et le néocortex affiche le plus grand nombre de neurones corticaux (environ 16 milliards) par rapport à la masse cérébrale, ainsi que par rapport à d’autres espèces de primates et de cétacés. Parmi les primates, le cortex humain contient de loin le plus grand nombre de neurones, avec environ deux fois plus de neurones que le cortex de chimpanzé et plus de dix fois plus que le cortex de macaque. Le plus frappant est que le cortex humain affiche également le nombre relatif et absolu le plus élevé de neurones pyramidaux des couches supérieures (couches II et III) par rapport aux autres espèces. Cette expansion des couches supérieures est déjà présente chez les singes et est encore élargie chez les Hominidés. Cette expansion est probablement importante dans l’évolution des circuits corticaux humains, car les neurones de la couche supérieure sont principalement impliqués dans les projections intracorticales. Leur nombre plus élevé contribue donc à des niveaux plus élevés de connectivité entre les zones corticales.
Des études utilisent actuellement des comparaisons d’organoïdes corticaux produits à partir de cellules souches humaines et des cellules souches de singe pour comprendre les origines développementales des différences spécifiques à la lignée humaine. Le développement du cortex humain est plus lent mais la densité en dendrites des neurones produits est plus importante. Des études complémentaires ont montré que cela est sans doute dû à la présence de 4 gènes SRGAP2 dans la lignée humaine au lieu d’un seul chez les singes proches (Schmidt et al., 2019). Egalement, les gènes Notch2NL, issus de duplications génétiques de Notch2 spécifiques à l’homme, augmentent le nombre de progéniteurs corticaux et augmentent la production neuronale lorsqu’ils sont surexprimés au cours du développement du cortex chez la souris (Fiddes et al., 2018).
Le développement des cellules gliales
Les cellules gliales du système nerveux ont des rôles largement plus étendus que celui de simple « colle » (en anglais : glue) qui leur a été assignées initialement. Elles jouent un rôle fondamental dans le développement et la physiologie du tissu nerveux.
Les cellules souches neuronales corticales donnent naissance tant aux neurones, qu’aux oligodendrocytes (qui forment les gaines de myéline dans le système nerveux central), et aux astrocytes. Attention, les cellules microgliales qui ont des fonction immunitaires proviennent de cellules hématopoiétiques embryonnaires qui sont rentrés dans le tissu neural avant la formation de la barrière hémato-encéphalique et ne proviennent pas des cellules souches neurales.
Les cultures des cellules souches neurales montrent qu’elles possèdent une horloge interne qui contrôle le temps pendant lequel elles produisent des progéniteurs de neurones, puis des progéniteurs de cellules gliales (Qian et al., 2000; Shen et al., 2006). Ainsi, chez la souris, en moyenne, une cellule de la glie radiaire se divise asymétriquement 8 à 9 fois pour former des précurseurs neuronaux puis environ 1/6ème de ces cellules de la glie radiaire se mettent à produire des cellules gliales (Beattie et al., 2017). Les autres meurent ou se différencient.
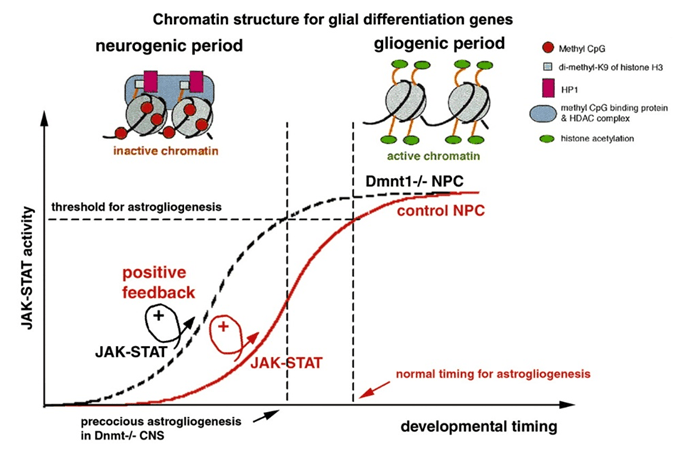
Des cellules souches neurales avec une triple potentialité puis une bipotence transitoire ont pu être cultivées : NA (neurones + astrocytes), NO (neurones + oligodendrocytes) et AO (astrocytes-oligodendrocytes). Des ligands orientant vers l’une ou l’autre voie ont été identifiés. Par exemple, le CNTF oriente fortement le destin des progéniteurs générés vers la voie astrocytaire et stimule également la prolifération des progéniteurs astrocytaires (Ravin et al., 2008). CNTF active le facteur de transcription STAT3 qui se fixe directement sur les promoteurs des gènes GFAP et S100, qui sont des gènes exprimés dans les cellules gliales. Dans les progéniteurs générés tôt dans le développement, l’ADN du promoteur de GFAP est méthylé, ainsi STAT3 ne peut pas déterminer les cellules en glie et ce sont des neurones qui sont d’abord produits avant que cette marque épigénétique inhibitrice soit enlevée (Fan et al., 2005). L’absence de l’ADN méthyltransférase de maintenance DNMT1 dans les précurseurs neuronaux aboutit à leur différenciation en astrocytes. A l’inverse, l’absence d’enzymes de déméthylation TET inhibe le potentiel des cellules souches neurales de donner des cellules gliales (MacArthur et al., 2024).
Le timing du passage de la production de neurones à la production de cellules gliales est donc étroitement régulé et a lieu 7 jours après la naissance chez la souris et 6 mois après la naissance chez les humains. Ce changement est dépendant aux facteurs évoqués ci-dessus mais aussi à l’activation de l’expression du gène NFIA qui code un facteur de transcription. Lors de la différenciation de cellules pluripotentes humaines (cellules ES ou iPS), l’expression transitoire de NFIA d’origine exogène accélère rapidement la production de cellules gliales (Tchieu et al., 2019).
Lors de la transition de la fin de la neurogenèse au début de la gliogenèse, les complexes répresseurs transcriptionnels Polycomb PRC1 et PRC2 sont nécessaires pour favoriser l’astrogliogenèse. La suppression des sous-unités centrales des complexes Polycomb : Ring1B, Ezh2 ou Eed dans les progéniteurs corticaux de souris prolonge la neurogenèse et retarde l’entrée dans la phase astrogénique. Notamment, les protéines du complexe Polycomb répriment l’expression du facteur de transcription neurogénine 1 (Ngn1), un suppresseur connu de l’astrogliogenèse. La mise sous silence de l’expression de Ngn1 est une condition préalable essentielle à la transition des progéniteurs neuraux vers le destin glial (Sun et al., 2001).
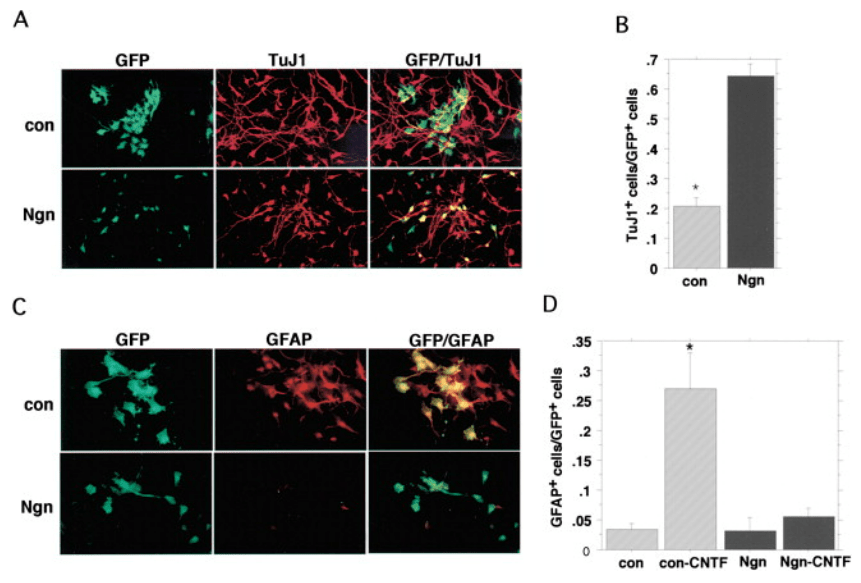
Des cultures primaires de cortex de rat au stade E14 ont été infectées par des rétrovirus permettant d’exprimer Ngn1 ou un gène contrôle (con). Quatre jours après mise en culture/infection, les cellules ont été fixées et soumises à une immunofluorescence pour les marqueurs de différenciation neuronale (TuJ1) et astrocytaire (GFAP). Les cellules infectées étaient positives pour la GFP (A et C). En (C), mais pas en (A), les cellules ont été traitées avec du CNTF pendant quatre jours pour favoriser la différenciation des astrocytes. Source : https://www.sciencedirect.com/science/article/pii/S0092867401002240
Signalons que la maturation du cortex est très lente notamment chez l’Homme et que la myélinisation, qui permet d’accélérer la conduction nerveuse au moins 20 fois, se développe jusqu’à la fin de l’adolescence pour certaines régions corticales. Le maximum de synapses est atteint vers 5 ans et le nombre de synapses décroit ensuite (Liu et al., 2012).
Régionalisation au sein du cortex
Il existe des différences régionales au sein du cortex qui sont fonctionnellement importantes car les différentes aires générées ont des connectivités et des fonctions spécifiques. Les différences peuvent même s’observer à l’échelle de la cytoarchitecture. Par exemple, les régions consacrées au traitement de l’information sensorielle, comme le cortex visuel, ont un nombre relativement important de cellules de la couche IV, qui forment la couche d’entrée d’informations tandis que les régions importantes pour la « sortie » de l’information du cortex, comme le cortex moteur primaire, ont un grand nombre de neurones pyramidaux de la couche V et relativement peu de cellules de la couche IV.
La spécification des différentes régions du cortex est sous le contrôle d’une cascade de régulations géniques avec à leur tête des gradients de morphogène comme les FGF.
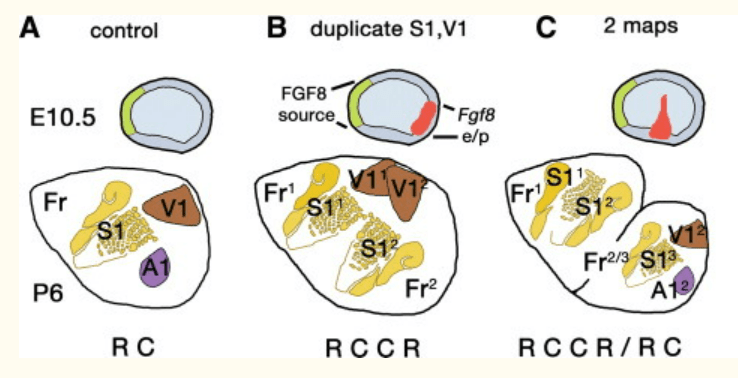
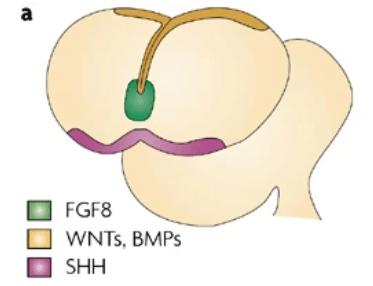
Les FGF sont produits dans la région la plus antérieure du système nerveux qu’on appelle l’ANR (pour Anterior Neural Ridge) (Eagleson et al., 2002). Cette région est induite chez la souris par l’endoderme viscéral antérieur (AVE), une structure extraembryonnaire qui produit aussi des FGF. A partir de l’ANR, FGF diffuse dans le futur cortex induisant des structures antérieures à forte concentration et des structures postérieures à faible concentration.
Ces gradients de morphogène se traduisent ensuite en gradients d’expression de facteurs de transcription : un gradient de Pax6 qui spécifie plutôt les régions antérieures et un gradient de Emx2 qui spécifie plutôt les régions postérieures. Les souris mutantes Pax6-/- présentent une hypertrophie des régions corticales postérieures au détriment des régions antérieures alors que c’est l’inverse pour les souris mutantes Emx2-/- (Muzio et Mallamaci, 2003). Cela démontre une inhibition croisée entre Pax6 et Emx2.
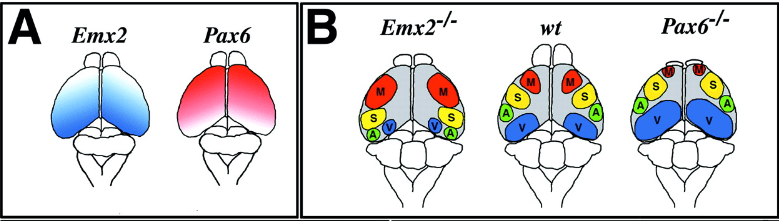
le cortex cérébral postérieur, puis son expression diminue progressivement vers le pôle cortical antérieur. Pax6 a un patron d’expression complémentaire.
Chez l’animal de type sauvage (wt), le cortex moteur (M) est principalement situé dans le cortex antérieur, et les autres régions sensorielles somatosensorielle (S), auditive (A) et visuelle (V) sont situées dans le cortex moyen et postérieur, respectivement. Chez les souris déficientes en Emx2, la répartition est décalés vers la région postérieure et une plus grande région est occupée par le cortex moteur. En revanche, chez les souris déficientes en Pax6, le cortex visuel est élargi et le cortex moteur est très réduit. Source : https://academic.oup.com/cercor/article/13/6/641/360937
Des études de transcriptomique et de ChiPseq ont permis de révéler les réseaux de régulation génétiques à l’oeuvre dans les cellules souches neurales de différentes régions du cortex (Ypsilanti et al., 2021).
Mise en place, maintien et modifications des connexions
Une fois les neurones générés et leur identité précisée, l’arborisation dendritique et la croissance axonale se mettent en place.
VOIR LA PAGE SUR LA CROISSANCE ET LE GUIDAGE AXONAL
Une proportion importante de progéniteurs neuronaux (les estimations varient entre 20 et 60%) meurt par apoptose durant le développement embryonnaire. Des souris n’exprimant pas Apaf-1 ou la caspase 9 qui sont des activateurs de l’apoptose présentent des zones progénitrices élargies (Yoshida et al., 1998). Dans les premiers jours après la naissance, des neurones en train d’être incorporés dans les circuits meurent également (autour de 30% des neurones). Le taux d’apoptose varie selon les régions corticales, avec plus de mort cellulaire dans le cortex moteur que dans le cortex somatosensoriel par exemple (Blanquie et al., 2017).
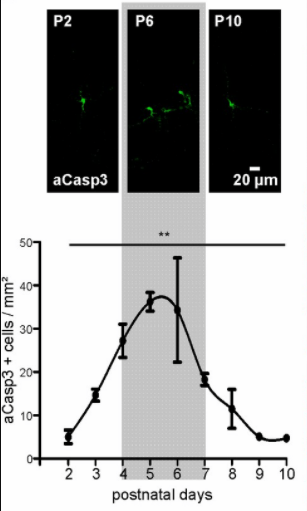
Ensuite, durant toute la vie post-embryonnaire, ce sont les synapses qui se font et se défont au gré des stimuli nerveux. Ces mécanismes contribuent à la plasticité du système nerveux. De manière plus subtile, des synapses peuvent se renforcer ou au contraire s’affaiblir.
Un exemple bien étudié d’affinement post-natal des connexions neuronales est constitué par les colonnes de dominance oculaire de la couche IV du cortex visuel. Les axones qui arrivent dans cette couche du cortex et qui apportent les informations en provenance de l’un ou de l’autre œil sont mélangés à la naissance. Progressivement, ces axones se séparent en territoires précis. Des expériences de suture d’un seul œil après la naissance chez le chat ou le singe ont montré que ce sont les stimuli en provenance de la rétine qui sont légèrement différents d’un œil à l’autre qui territorialisent les projections. Quand un œil est suturé, les projections axonales qui transmettent ces informations au cortex se réduisent tandis que celles en provenance de l’autre œil sont maintenues donc finissent par être relativement plus grandes. Ces expériences ont aussi mis en évidence l’existence d’une période critique. Ces réarrangements de projection axonale et de synapse n’ont lieu de manière significative que sur une période assez courte. Après, des réarrangements peuvent avoir lieu mais sur une échelle plus petite. Au niveau moléculaire, les réarrangements impliquent des sécrétions différentielles de facteurs neurotrophiques tels le BDNF et des expressions différentielles de molécules impliquées dans l’adhérence ou la communication juxtacrine telle les éphrines. Dans le cas du BDNF, il a été bien montré que la quantité de BDNF qu’un neurone est capable de produire et de sécréter détermine la densité de ses épines dendritiques et qu’il joue un rôle dans la compétition entre les neurones du cortex visuel pour se connecter (English et al., 2012).
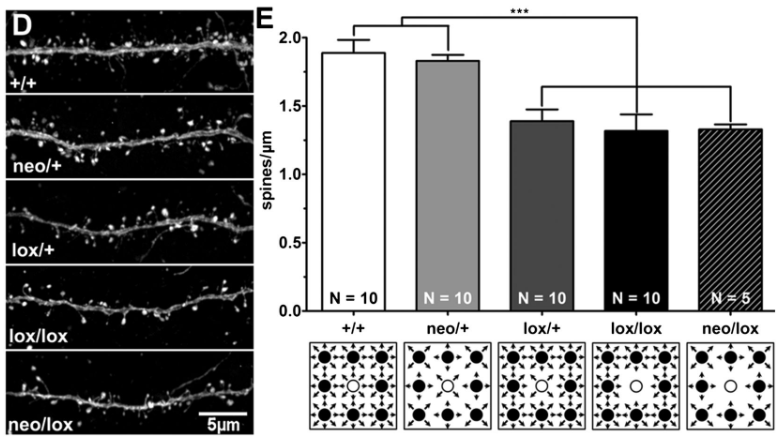
A un niveau d’intégration inférieur, le nombre de synapses connectées à un neurone peut changer, ce qui se traduit par une augmentation du nombre de ses épines dendritiques, c’est-à-dire les éléments post-synaptiques. La force des synapses peut être modulée ce qui peut se traduire morphologiquement par des changements de la forme des épines dendritiques. Au niveau moléculaire, par exemple, les quantités de récepteurs de neurotransmetteurs peuvent être modifiées.
Bien que des modifications aient lieu toute la vie, elles ralentissent fortement à partir de la puberté. Certaines régions du cerveau hors cortex restent néanmoins assez « plastiques », notamment l’hippocampe impliqué dans la mémorisation.
L’édition de la séquence des ARNm et tout particulièrement des transitions de A (adénosine) à I (inosine) est un phénomène courant pour des gènes impliqués dans le neurodéveloppement et la physiologie neuronale (Ekdahl et al., 2012: Mehler et al., 2007). Elle se produit au niveau d’adénosines isolées uniques ou sur de nombreuses adénosines voisines dans une région étendue et est catalysée par une famille d’adénosine désaminases appelée ADAR. L’édition de A à I dans les régions codantes pour les protéines peut conduire à substitutions d’acides aminés ou si elle a lieu dans les 3’UTR peut modifier la fixation de microARN. Ces modifications sont des contributeurs majeurs à la diversité globale des séquences d’ARN dans le cerveau humain et amplifient la fonctionnalité de nombreuses protéines exprimées dans les neurones, notamment celles impliquées dans la transmission synaptique et les voies de signalisation neuronaux. Elles régulent étroitement la perméabilité au Ca2+, et remodèlent le cytosquelette d’actine au niveau des synapses excitatrices, entre autres fonctions. Des milliers d’éditions A à I ont pu être caractérisés durant les phases de neurodéveloppement et leurs fonctions sont en cours d’investigation (Cuddleston et al., 2022).
Produire in vitro des neurones corticaux à partir de cellules souches
De nombreux protocoles existent pour différencier des neurones corticaux à partir de cellules ES ou iPS. Ils s’appuient sur la signalisation qui se déroule lors du développement embryonnaire normal : il s’agit d’abord d’induire du tissu neural (donc de bloquer la signalisation BMP pour mimer l’effet de l’organisateur de Spemann), puis d’induire la régionalisation antérieure. Selon les besoins, on peut manipuler la signalisation qui permet la régionalisation dorso-ventrale (par exemple stimuler la voie de signalisation Shh si on souhaite obtenir des interneurones inhibiteurs à GABA qui sont d’origine ventrale).
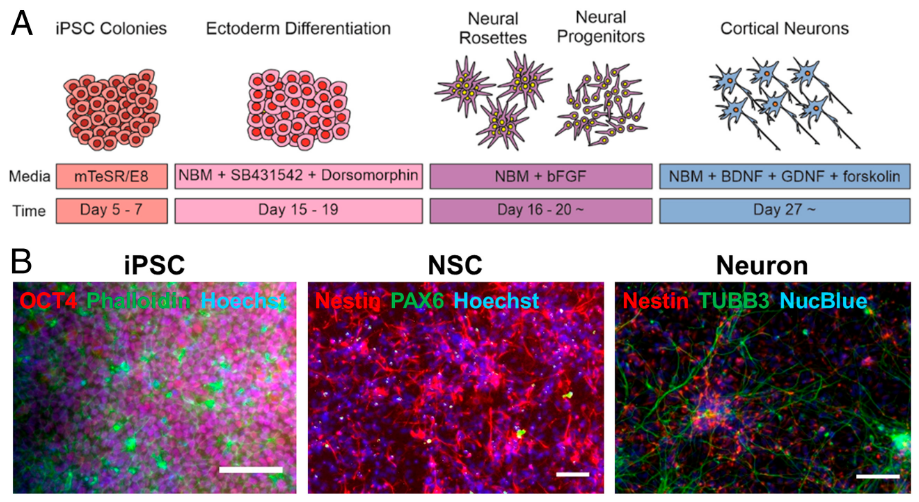
Les organoïdes sont des cultures agrégées auto-organisées dérivées de cellules souches (notamment humaines) qui génèrent plusieurs types de cellules et une structure reflétant un organe en développement. Des organoïdes corticaux ont pu être générés à partir de la fin des années 2010 avec des protocoles de plus en plus performants et leur développement suit remarquablement bien la séquence d’évènements qui ont lieu in vivo. Par exemple, les organoïdes corticaux sont d’abord composés de cellules progénitrices multipotentes précoces : cellules neuroépithéliales et cellules gliales radiaires (Subramanian et al. 2017). Les cellules progénitrices prolifèrent de manière robuste et utilisent des programmes de division qui sont très similaires au développement cortical endogène au niveau transcriptionnel (Pollen et al. 2019, Velasco et al. 2019). Des neurones excitateurs répartis en plusieurs couches et des astrocytes ont pu être obtenus (Velasco et al. 2019), de même que des oligodendrocytes qui ont formé des gaines de myéline autour des axones (Marton et al., 2019). Cependant, il existe des différences avec ce qu’il se passe in vivo. Par exemple, certains types de neurones excitateurs dérivés d’organoïdes ont une expression réduite de certains gènes marqueurs tel que SATB2 pour les neurones des couches supérieures. Or SATB2 est clairement une protéine importante vu que chez l’homme, les mutations du gène SATB2 provoquent un retard de développement, une déficience intellectuelle, de l’épilepsie et un développement de la parole absent ou limité (Zarate et al., 2016). Autre problème, la taille des organoïdes est limitée par une mauvaise oxygénation des régions profondes où il y a hypoxie et nécrose.
Ces outils sont étudiés pour comprendre l’origine de malformations ou de maladies cérébrales (comme par exemple le syndrome de Rett (Tenreiro et al., 2025)) mais aussi dans un objectif évo-dévo par exemple en comparant le développement d’organoïdes issus de cellules souches de chimpanzé, de gorille et d’humains (Pollen et al., 2019; Benito-Kwiecinski et al., 2021).
EN DIRECT DES LABOS :
Utilisation d’organoïdes de cerveau :
Présentation du laboratoire Génétique et développement du cortex cérébral (Institut Imagine – Paris) :
QUELQUES EQUIPES FRANCOPHONES QUI TRAVAILLENT SUR LE SUJET :
Effets des résidus médicamentaux sur le développement du cerveau – INRAE, Tours
Mécanismes physiologiques et pathologiques du développement cortical – IGBMC, Strasbourg
Génétique et développement du cortex cérébral – Institut IMAGINE, Paris
- Adhérences cellule-cellule
- Arabidopsis thaliana
- Axe antéro-postérieur chez la drosophile
- Biomécanique du développement
- Caenorhabditis elegans
- Concepts principaux
- Contrôle de la traduction
- Contrôle de la transcription
- Contrôle génétique et épigénétique
- Croissance du tube pollinique et double fécondation chez les Angiospermes
- Croissance et guidage axonal
- Des modèles animaux moins classiques
- Développement de l’oeil des Vertébrés
- Développement et évolution
- Et l’Humain ?
- Exercices sur l’ovogenèse, la spermatogenèse et la fécondation
- Exercices sur le contrôle de l’expression des gènes
- Exercices sur le développement des bourgeons de membre
- Exercices sur le développement des muscles striés squelettiques
- Exercices sur le développement des végétaux et les hormones végétales
- Exercices sur les cycles et les divisions cellulaires
- Exercices sur les étapes du développement, les inductions embryonnaires et la mise en place des axes de polarité
- Exercices sur les matrices extracellulaires, le cytosquelette et les adhérences cellule-cellule
- Exercices sur les voies de signalisation
- Glossaire
- Glossaire des termes liés à la génétique
- Glossaire des termes liés au cytosquelette, la matrice extracellulaire, l’adhérence et la migration cellulaire
- Hématopoïèse et développement des cellules du système immunitaire
- Histoire de la biologie cellulaire et de la biologie du développement
- L’acide rétinoïque
- L’apoptose
- L’autophagie
- L’organogenèse
- L’ovogénèse prépare le développement embryonnaire
- La drosophile
- La fécondation
- La formation des somites
- La gastrulation
- La gastrulation (version allégée)
- La métamorphose chez les Hexapodes et les Amphibiens
- La neurogénèse chez les mammifères adultes
- La neurulation
- La poule
- La signalisation calcique
- La souris
- La superfamille TGFβ et ses voies de signalisation
- La voie de signalisation de l’auxine et ses rôles
- La voie de signalisation Hedgehog
- La voie de signalisation Hippo et ses composants YAP/TAZ
- La voie de signalisation Notch
- Le clivage
- Le cytosquelette
- Le destin des cellules et les réseaux de régulation génique
- Le développement des bourgeons de membre
- Le développement des muscles striés squelettiques
- Le développement des organes génitaux et des cellules germinales
- Le méristème apical caulinaire en phase végétative et lors de la formation d’une fleur
- Le poisson zèbre
- Le xénope
- Les cellules des crêtes neurales
- Les cellules et les gènes en action dans le développement
- Les cellules souches
- Les cellules tumorales
- Les cycles et les divisions cellulaires
- Les étapes du développement
- Les étapes du développement embryonnaire d’Arabidopsis thaliana et leur contrôle
- Les inductions embryonnaires et les gradients de morphogène
- Les matrices extracellulaires animales
- Les organismes modèles
- Les outils pour étudier l’expression et la fonction des gènes
- Les parois des cellules végétales
- Les techniques et les outils pour la biologie cellulaire
- Les transitions épithélio-mésenchymateuses et les migrations cellulaires
- Les vésicules extracellulaires
- Les voies de signalisation
- Les voies de signalisation FGF
- Mise en place des axes chez les Vertébrés
- Structures et processus cellulaires
- Voies de signalisation WNT
