
Par Patrick Pla, Université Paris-Saclay

Les Amphibiens ont été utilisés depuis longtemps pour étudier les premières étapes du développement embryonnaire chez les vertébrés, car leurs ovocytes sont gros (1 à 2 mm de diamètre), nombreux (500 à 800 pour la ponte d’une femelle de xénope), faciles à manipuler et fécondés à l’extérieur de l’animal. Ainsi le développement de l’embryon précoce est facilement suivi. Les Amphibiens se prêtent particulièrement bien à une approche de criblage rapide et efficace de gènes candidats ou de traitements pharmacologiques.
Leur développement est rapide et sa progression peut être modulée par la température selon les besoins de l’expérimentateur (en évitant des changements trop violents cependant, notamment lors de la gastrulation car cela peut amener à des malformations).
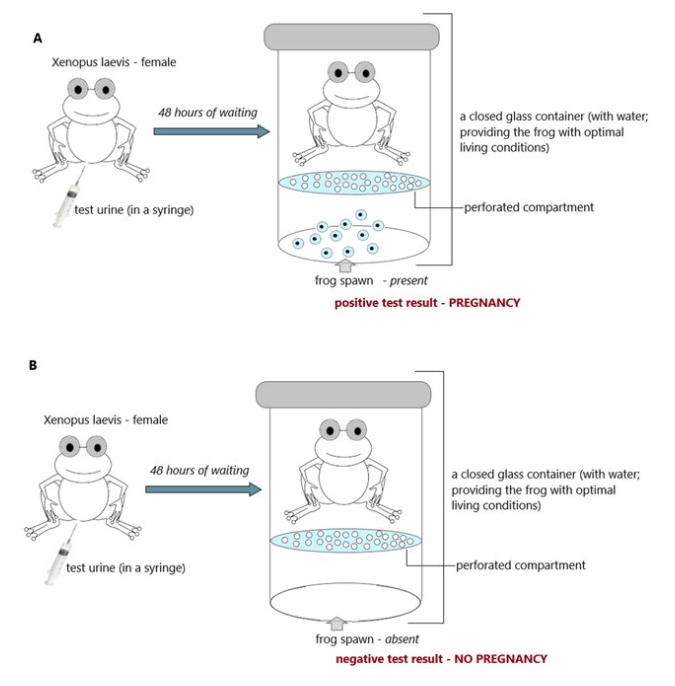
L’utilisation du xénope en particulier qui est originaire d’Afrique a été popularisé comme test de grossesse à partir des années 1930 et est ensuite devenu un modèle privilégié parmi les « grenouilles » pour l’étude du développement.

Vous pouvez découvrir le cycle du développement du xénope sur ce schéma interactif.
Des dessins d’observation de l’ensemble des stades de développement du xénope sont disponibles sur ce lien.
Le xénope est un modèle où on peut très facilement voir les mouvements de la gastrulation. Vous pouvez regarder cette vidéo de sa gastrulation et de sa neurulation.

Deux espèces de Xenopus sont utilisées dans la recherche sur le développement : X. laevis et X. tropicalis. Chaque espèce offre des avantages spécifiques. Xenopus laevis produit de gros embryons qui sont excellents pour la microchirurgie embryonnaire, l’analyse de la surexpression des gènes, les études biochimiques. Il continue d’être un organisme modèle important, même s’il est peu adapté à l’analyse génétique à cause de son allotétraploïdie (voir ce site pour comprendre son histoire génétique).


De son côté, Xenopus tropicalis est un organisme diploïde qui facilite les études de perte de fonction. Les ovocytes et les embryons de Xenopus tropicalis sont cependant plus petits que ceux de Xenopus laevis. Le génome des xénopes a une conservation importante avec l’Homme. Celui de Xenopus tropicalis contient des orthologues pour environ 79 % des gènes identifiés dans les maladies humaines. De plus, son génome a un degré élevé de synténie avec le génome humain, contenant de longues régions équivalentes avec des gènes positionnés dans un ordre conservé.
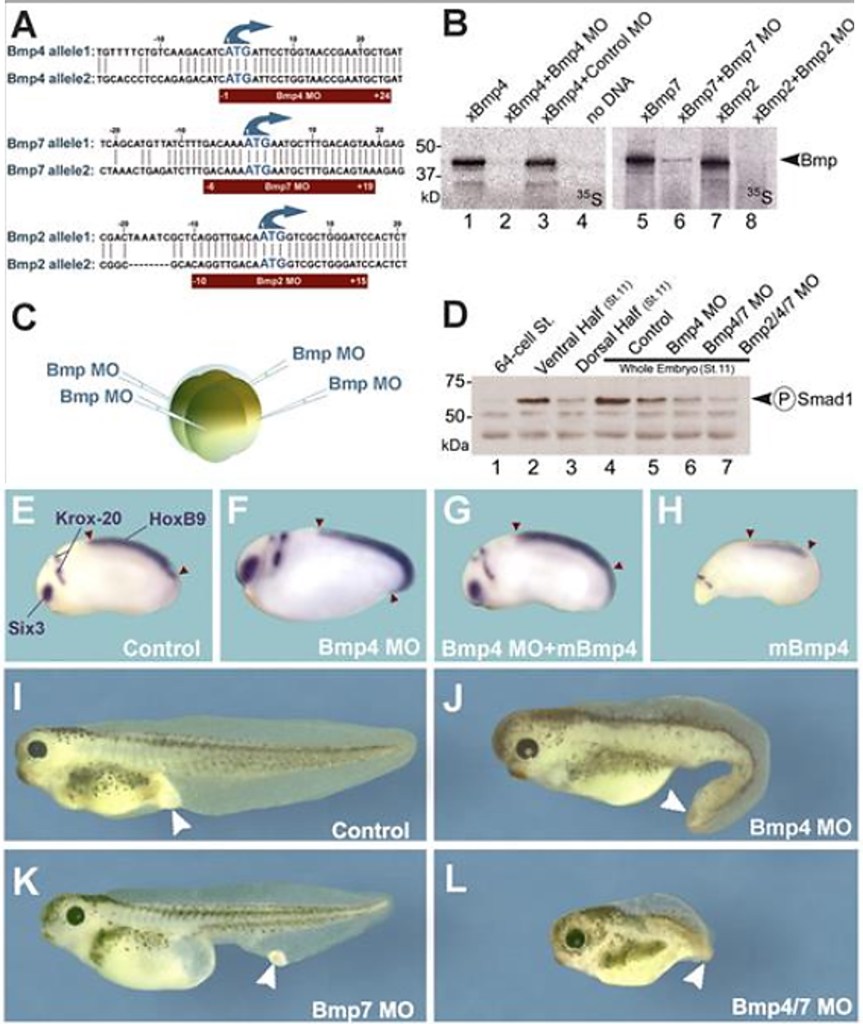
L’expression des gènes dans les embryons de xénope peut être facilement manipulée à l’aide de microinjections d’ARNm et d’oligonucléotides morpholino antisens (MO) pour un gain ou une perte de fonction, respectivement.

Les morpholino antisens (MO) sont conçus pour cibler soit le site de départ de la traduction (le codon d’intiation AUG), soit un site d’épissage du transcrit primaire de l’ARNm cible.


Source : https://www.sciencedirect.com/science/article/pii/S0012160613006568
Voici un autre exemple d’utilisation de MO pour étudier la fonction des morphogènes de la famille BMP :
Un autre avantage majeur de l’utilisation de Xenopus pour étudier le développement est la capacité de générer des animaux transgéniques. L’incorporation d’ADN exogène dans le génome d’un zygote a été développée chez Xenopus tropicalis et Xenopus laevis à la fin des années 1990. Lorsqu’il correspond à un rapporteur fluorescent comme la GFP sous le contrôle d’un promoteur spécifique, l’expression d’un transgène peut être surveillée au fil du temps dans un embryon vivant en développement. Des lignées rapportrices, par exemple de l’activité d’une voie de signalisation, peuvent être étudiées. Une activation transgénique inductible peut en plus fournir une expression spatio-temporelle contrôlée.
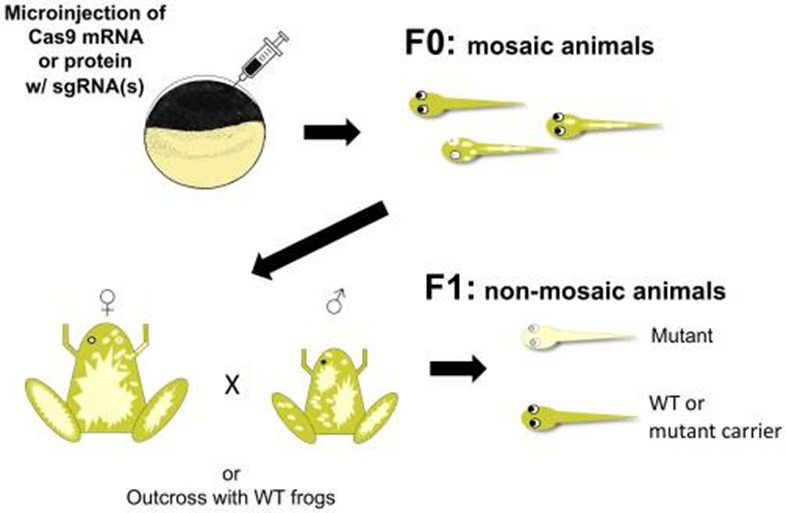
La technique CRISPR/Cas9 peut être utilisée pour créer des mutations gain ou perte-de-fonction ou générer des lignées rapportrices.
En plus des méthodes perte- et gain-de-fonction efficaces et peu coûteuses, Xenopus présente des avantages pour l’analyse comparative des structures. En effet, l’injection d’une seule cellule de l’embryon à deux cellules peut cibler le côté gauche ou droit de l’embryon. En utilisant des traceurs fluorescents (GFP) ou enzymatiques (β-galactosidase), on peut facilement détecter le côté injecté de l’embryon et le comparer au côté non injecté. De telles manipulations sont très utiles car elles fournissent un contrôle interne chez le même animal pour chaque expérience. En plus de l’injection au stade 2 cellules, les embryons de xénope ont une carte du destin cellulaire bien définie pour chaque système organique. Cela permet des injections ciblées, où l’expression des gènes peut être modifiée dans des organes ou des tissus spécifiques.
Les embryons de xénope et notamment de Xenopus laevis qui sont plus gros se prêtent très bien à des expériences de microchirurgie embryonnaire. Fait important, parce que les premiers stades de clivage sont holoblastiques (par opposition aux clivages méroblastiques chez les embryons de poule ou de poisson zèbre), le vitellus (réserves énergétiques) est distribué à toutes les cellules, de sorte que les explants ne nécessitent aucune nutrition supplémentaire car ils peuvent survivre en utilisant les nutriments stockés dans les cellules. Le tissu disséqué se développe et se différencie dans une simple solution saline. Le xénope est aussi utile pour produire des cultures cellulaires de cellules pluripotentes : les cellules de la calotte (ou coiffe) animale, prélevées sur des blastulas. Sans intervention, ces cellules finissent par se développer en petits organoïdes épidermiques. Mais en présence de différents ligands et avec des combinaisons d’expression de gènes (induites par des injections d’ARNm dans l’embryon précoce au préalable), ces cellules peuvent aussi donner des tissus neuraux, des cellules de crêtes neurales, des cellules mésodermiques ou endodermiques.
Le xénope n’a pas été que utile pour étudier le développement embryonnaire stricto sensu mais a aussi permis des avancées dans des domaines de génétique et de biologie cellulaire plus fondamentaux. Par exemple, le premier gène Eucaryote séquencé à été l’ADN codant l’ARN ribosomal 5S du xénope en 1968 et des éléments importants contrôlant le cycle cellulaire ont été mis en évidence chez le xénope avant d’être découverts dans d’autres modèles.

Si on réalise un transfert cytoplasmique d’un ovocyte mature non fécondé vers l’une des cellules d’un embryon à 2 cellules, cette cellule ne se divise plus (elle est dit « en arrêt CSF ») démontrant la capacité du cytoplasme de l’ovocyte mature à maintenir l’activité MPF et à bloquer le cycle. Source
SITE DE REFERENCE : Xenbase
EN DIRECT DES LABOS :
QUELQUES EQUIPES DE RECHERCHE FRANCOPHONES QUI TRAVAILLENT SUR LE SUJET :
Equipe « Cellules souches et neurogenèse dans la rétine » – NeuroPSI, Université Paris-Saclay
Plateforme Xénope de l’U1035 INSERM – Biothérapie des maladies génétiques inflammatoires et cancer – Université de Bordeaux
- Adhérences cellule-cellule
- Arabidopsis thaliana
- Axe antéro-postérieur chez la drosophile
- Biomécanique du développement
- Caenorhabditis elegans
- Concepts principaux
- Contrôle de la traduction
- Contrôle de la transcription
- Contrôle génétique et épigénétique
- Croissance du tube pollinique et double fécondation chez les Angiospermes
- Croissance et guidage axonal
- Des modèles animaux moins classiques
- Développement de l’oeil des Vertébrés
- Développement et évolution
- Et l’Humain ?
- Exercices sur l’ovogenèse, la spermatogenèse et la fécondation
- Exercices sur le contrôle de l’expression des gènes
- Exercices sur le développement des bourgeons de membre
- Exercices sur le développement des muscles striés squelettiques
- Exercices sur le développement des végétaux et les hormones végétales
- Exercices sur les cycles et les divisions cellulaires
- Exercices sur les étapes du développement, les inductions embryonnaires et la mise en place des axes de polarité
- Exercices sur les matrices extracellulaires, le cytosquelette et les adhérences cellule-cellule
- Exercices sur les voies de signalisation
- Glossaire
- Glossaire des termes liés à la génétique
- Glossaire des termes liés au cytosquelette, la matrice extracellulaire, l’adhérence et la migration cellulaire
- Hématopoïèse et développement des cellules du système immunitaire
- Histoire de la biologie cellulaire et de la biologie du développement
- L’acide rétinoïque
- L’apoptose
- L’autophagie
- L’organogenèse
- L’ovogénèse prépare le développement embryonnaire
- La drosophile
- La fécondation
- La formation des somites
- La gastrulation
- La gastrulation (version allégée)
- La métamorphose chez les Hexapodes et les Amphibiens
- La neurogénèse chez les mammifères adultes
- La neurulation
- La poule
- La signalisation calcique
- La souris
- La superfamille TGFβ et ses voies de signalisation
- La voie de signalisation de l’auxine et ses rôles
- La voie de signalisation Hedgehog
- La voie de signalisation Hippo et ses composants YAP/TAZ
- La voie de signalisation Notch
- Le clivage
- Le cytosquelette
- Le destin des cellules et les réseaux de régulation génique
- Le développement des bourgeons de membre
- Le développement des muscles striés squelettiques
- Le développement des organes génitaux et des cellules germinales
- Le développement du cortex
- Le méristème apical caulinaire en phase végétative et lors de la formation d’une fleur
- Le poisson zèbre
- Les cellules des crêtes neurales
- Les cellules et les gènes en action dans le développement
- Les cellules souches
- Les cellules tumorales
- Les cycles et les divisions cellulaires
- Les étapes du développement
- Les étapes du développement embryonnaire d’Arabidopsis thaliana et leur contrôle
- Les inductions embryonnaires et les gradients de morphogène
- Les matrices extracellulaires animales
- Les organismes modèles
- Les outils pour étudier l’expression et la fonction des gènes
- Les parois des cellules végétales
- Les techniques et les outils pour la biologie cellulaire
- Les transitions épithélio-mésenchymateuses et les migrations cellulaires
- Les vésicules extracellulaires
- Les voies de signalisation
- Les voies de signalisation FGF
- Mise en place des axes chez les Vertébrés
- Structures et processus cellulaires
- Voies de signalisation WNT
