
Par Patrick Pla, Université Paris-Saclay
FGF veut dire facteur de croissance des fibroblastes. Le terme de facteur de croissance est assez limitant, et même si ces facteurs font effectivement croitre les populations de cellules par prolifération, leurs rôles ne se limitent pas à cela. Les facteurs de croissance peuvent aussi jouer un rôle instructeur qui provoque des changements sur la détermination des cellules et non pas seulement un rôle sur leur quantité. Et, comme nous allons le constater, les FGF ont des cibles nettement plus diversifiées que les fibroblastes !
La famille FGF est composée de 22 membres (FGF 1-14 et 16-23). Chacun possède un domaine central conservé de 120 acides aminés. Ces ligands sont organisés en sept groupes basés sur des relations phylogénétiques.
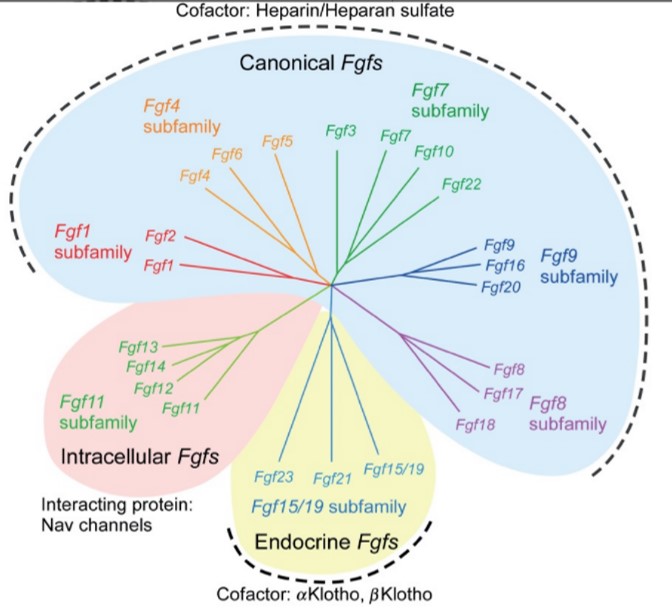
Les FGF se lient avec une faible affinité aux protéoglycanes d’héparane sulfate (HSPG) présents sur la plupart des surfaces cellulaires et des matrices extracellulaires. Cette interaction est nécessaire à leur activité.
Les premiers FGF à avoir été identifiés sont les FGF1 et FGF2, purifiés à partir d’extraits d’adénohypophyse et montrant des propriétés mitogéniques (Armelin, 1973; Gospodarowicz et al., 1974). Il existe deux FGFs chez C. elegans : let-756 (let, lethal) essentiel au développement du nématode, et egl-17 (egl, egg-laying defective) requis pour la migration des myoblastes sexuels
(Burdine et al., 1997). Egl-17 est clairement l’orthologue de la famille FGF11/12/13/14 des Vertébrés (Itoh, 2007). Les relations phylogénétiques de let-756 sont moins définies. Trois homologues ont été identifiés chez D. melanogaster : branchless, orthologue de la plupart des FGF des Vertébrés et qui est impliqué dans la migration cellulaire et la ramification des trachées, et thisbe et pyramus impliqués dans le développement (Stathopoulos et al., 2004; Sutherland et al., 1996). Heartless est le nom de leur récepteur chez la drosophile.

Au cours de l’évolution, des évènements supplémentaires de duplication du génome chez le poisson-zèbre portent le nombre de FGFs à 27 comprenant 21 homologues humains (tous excepté le FGF9), et 6 paralogues (Itoh, 2007).
Les FGFs agissent pour la plupart via leurs récepteurs FGFRs. Selon la combinaison ligand/récepteur, différentes voies de signalisation sont activées et interviennent dans une multitude de processus comme l’angiogenèse, la prolifération et la survie cellulaires, la différenciation ou encore la migration. Chez les mammifères, quatre gènes codant des récepteurs de FGF (FGFR1–4) ont été identifiés mais la diversité est bien plus grande à cause de multiples épissages alternatifs (Ornitz et Itoh, 2015; Brewer et al., 2016).

Du fait de cette implication dans des processus fondamentaux, de nombreuses pathologies sont associées à des niveaux protéiques anormaux ou à des mutations des FGFs ou de leurs récepteurs. Ils sont notamment souvent associés à plusieurs types de cancer ou à des défauts de développement (Ornitz and Itoh, 2015).
De nombreux FGF présentent des interactions de haute affinité avec plusieurs FGFR, tandis que certains activent des récepteurs uniques ou des isoformes de récepteurs. La plupart des FGF ont démontré une activité mitogène dans une variété de systèmes. Cependant, quelques-uns, les FGF 11-14, 19 et 21-23, ne présentent pas de fonctions prolifératives. La capacité mitogène d’un FGF est probablement dépendante du ou des FGFR avec lesquels il interagit. Par exemple, le FGF-19 joue un rôle dans la régulation de la synthèse du cholestérol et des acides biliaires grâce à une interaction unique non héparine-dépendante avec le FGFR4. Quant à FGF23, il joue un rôle dans la régulation physiologique du métabolisme du phosphate et de la vitamine D.
Les récepteurs aux FGF sont des récepteurs tyrosine kinase. Les modèles classiques d’activation du récepteur tyrosine kinase impliquent la liaison du ligand, la dimérisation du récepteur, la transactivation du domaine tyrosine kinase, la phosphorylation des tyrosines intracellulaires. Ces sites phosphorylés permettent l’ancrage et l’activation des effecteurs (via leur domaine SH2 ou PTB) qui orchestrent à leur tour l’activation des voies de signalisation en aval (Lemmon et Schlessinger 2010).

Différents seuils de force et de stabilité des dimères peuvent également entrer en jeu, ainsi que la dynamique de signalisation engagée par chaque voie en aval (Vasudevan et al. 2015; Zinkle et Mohammadi 2018; Li et Elowitz 2019). Pour les FGFR, où les voies en aval ont été particulièrement bien étudiées, ERK1/2 et Akt sont les principaux effecteurs (Lanner et Rossant 2010; Brewer et al., 2016), mais la voie STAT peut également être activée ainsi que la voie PLCγ.

Source : https://elifesciences.org/articles/07186
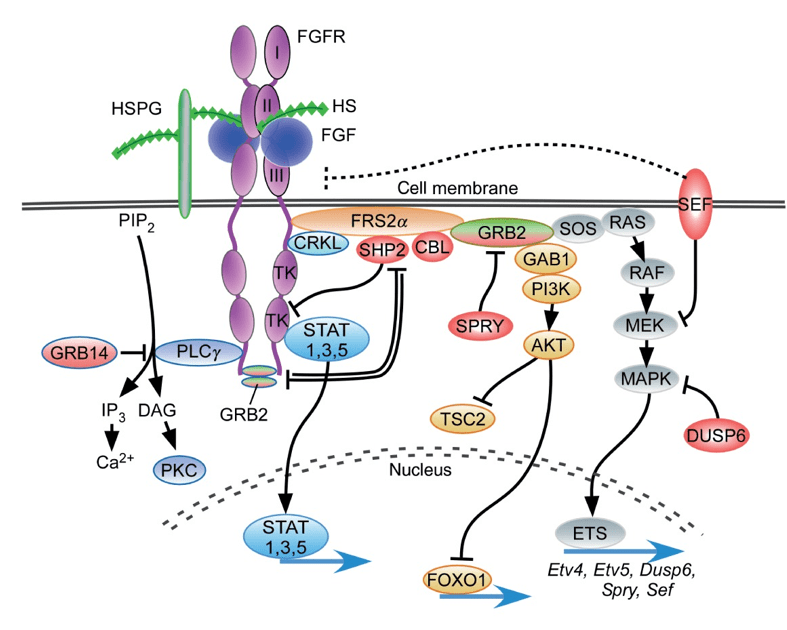

Une version commentée de ce schéma est disponible en vidéo.
D’après https://www.frontiersin.org/articles/10.3389/fnmol.2019.00121/full
Les récepteurs FGFR peuvent aussi être endocytés à la suite de l’activation de leur voie de signalisation en aval. Les récepteurs peuvent alors être dégradés ou recyclés à la membrane plasmique. L’endocytose et les mécanismes de trafic intracellulaire ont une influence importante sur la dynamique spatiale et temporelle du signal, qui influence les effets biologiques d’une exposition aux FGF (Sorkin et von Zastrow, 2009). La mutation perte-de-fonction de LIS1 qui code une protéine associée à la dynéine et au trafic vésiculaire aboutit à un phénotype rappelant une déficience en signalisation FGF (comme par exemple des membres tronqués voire absents dûs à l’absence de réponse en provenance de l’AER produisant des FGF au cours de la croissance des bourgeons de membre).

Avec d’autres expériences, Liu et al. (2018) ont montré que LIS1 était essentiel pour ramener des FGFR à la membrane plasmique après endocytose activée par le ligand. En son absence, les FGFR sont trop envoyés dans les lysosomes et sont dégradés de manière excessive, rendant moins efficace la réception du signal FGF.
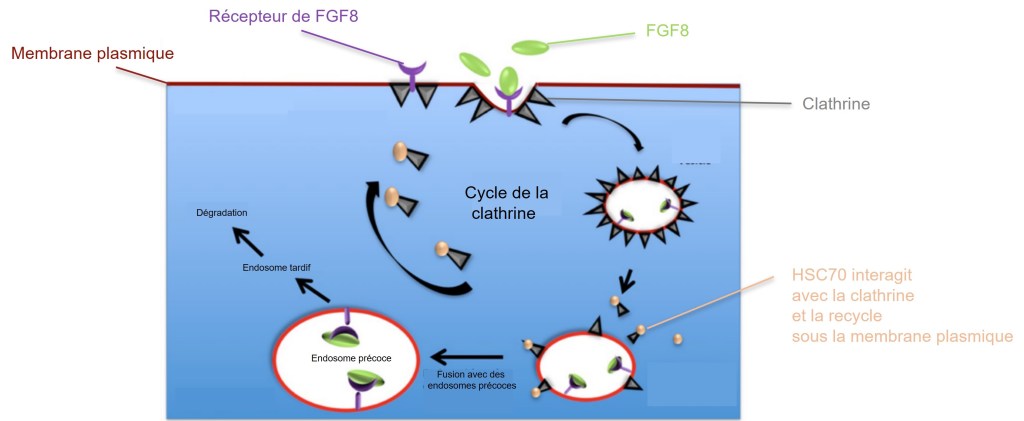
Cependant, une destruction des récepteurs et de leurs ligands est quelque fois souhaitable. Par exemple, lors de la gastrulation du poisson-zèbre, le gradient FGF8 est trop étendu si les récepteurs liés à FGF8 ne sont pas suffisamment dégradés. Les récepteurs liés à leur ligand sont d’abord endocytés dans des vésicules couvertes de clathrine. Une protéine appelée HSC70 est essentielle pour désassembler la clathrine de ces vésicules avant que celles-ci ne fusionnent avec les endosomes. Si il y a une mutation perte-de-fonction du gène codant HSC70 alors il n’y a plus suffisamment de clathrine disponible pour les endocytoses et le gradient de FGF8 s’étend causant des défauts de développement (Rengarajan et al., 2014).
Parmi les cibles de ERK activés par la signalisation FGF on peut signaler DNMT3A, une DNA-méthyltransférase qui méthyle l’ADN de novo sur des cytosines (ajout de nouvelles méthylations et non simple entretien) aboutissant à une inhibition de la transcription. La phosphorylation de DNMT3A par ERK empêche la DNA-méthyltransférase d’agir et d’inhiber la transcription.

(A) Schéma du DNMT3A humain montrant les sites de phosphorylation (S255) et d’amarrage d’ERK (ERK D-domain). (B) Les sites de phosphorylation et d’amarrage d’ERK1/ERK2 sont conservés sur DNMT3A chez les vertébrés (Ec, Equus caballus ; Gg, Gallus gallus ; Hs, Homo sapiens ; Mm, Mus musculus ; Rn, Rattus norvegicus). (C) Test de l’activité kinase ERK2 (en utilisant du γ-32P-ATP) sur des formes sauvages ou mutantes de GST-hDNMT3A (GST est un tag pour mieux reconnaitre et isoler la protéine). Il y a des mutations soit sur le site de phosphorylation S255, soit sur les sites d’amarrage de ERK. (D) Etude par western-blot utilisant des anticorps anti-DNMT3A-phosphoS255 (qui reconnait la forme phosphorylée) ou anti-DNMT3A (qui reconnait toutes les formes) après phosphorylation in vitro par ERK2 de formes sauvages ou mutantes de GST-hDNMT3A. (E) Les bourgeons de membre (isolés à partir d’embryons de poulet au jour 4) ont été séparés en régions proximale (P), moyenne (M) et distale (D). Des quantités égales de protéines de chaque région de bourgeon de membre ont été analysées par western-blot avec des anticorps anti-DNMT3A-phosphoS220 (le site de phosphorylation chez le poulet est décalé par rapport à l’humain) anti-DNMT3A, anti-phosphoERK, anti-ERK ou anti-β-actine. On constate que plus on est proche de la partie distale du bourgeon de membre et de l’AER qui produit FGF, plus ERK et DNMT3A sont phosphorylés. Source : https://www.cell.com/cell-reports/fulltext/S2211-1247(14)00622-6
Dans le bourgeon de membre, les FGF en provenance de la partie distale (AER) inhibent DNMT3A via la phosphorylation par ERK, empêchant d’inhiber la transcription de Sox9 par méthylation de l’ADN. Le maintien de l’expression de Sox9 est crucial dans le mésenchyme pour qu’il puisse encore produire du cartilage durant toute la croissance du bourgeon de membre.
Alors que dans la plupart des types cellulaires, le FGF induit la prolifération et
protège de l’apoptose, dans les chondroblastes, il provoque un arrêt de la croissance et contribue à leur différenciation en chondrocytes. Toutes les protéines Rb deviennent déphosphorylées lors du traitement par FGF et l’activité du complexe cycline E/CDK2 (qui contrôle la progression durant la phase G1) est inhibée (Kalupoeva et Basilico, 2012). L’expression de p21 est augmentée. C’est exactement l’inverse de ce que l’on observe dans la plupart des cellules, ce qui montre bien que l’action des ligands sur leurs cibles est contexte-dépendant et dépend de la compétence des cellules.
Lors de cette fonction inhibitrice de la prolifération des chondroblastes et activatrice de la différenciation en chondrocytes, FGF agit via le récepteur FGFR3. Ce récepteur est constitutivement actif à cause de mutations chez des patients atteints de dysplasie (ou nanisme) thanatophore, qui présentent des os longs et des côtes raccourcis (leur croissance dépend initialement du cartilage). La forme mutée de FGFR3 s’autophosphoryle sur les tyrosines intracellulaires et active la voie STAT en aval de ce récepteur sans qu’il y ait de ligands FGF et aboutit à la différenciation de chondrocytes alors même que le nombre de précurseurs n’est pas suffisant pour aboutir à une taille normale du squelette (Webster et Donoghue, 1996).

Source : https://www.embopress.org/doi/epdf/10.1002/j.1460-2075.1996.tb00384.x
Implication de la signalisation FGF dans le développement précoce de la souris
Dans le blastocyste de souris à E4.5, les populations Nanog+ (épiblaste) et Gata6+ (endoderme primitif) sont ségrégées par la signalisation FGF4 (Yamanaka et al., 2010). En effet, les cellules exprimant le récepteur FGFR1 ainsi que la signalisation autocrine FGF4 s’engagent dans l’épiblaste, tandis que la signalisation paracrine FGF4 via FGFR1 et FGFR2 détermine les cellules de l’endoderme primitif.

Les niveaux d’ERK relativement faibles dans les cellules FGFR1+ déclenchent une rétroaction négative, augmentant l’expression de Nanog et l’identité pluripotente de l’épiblaste. En revanche, la signalisation FGF-ERK dans les cellules FGFR1+/FGFR2+ active des cibles alternatives, telles que DUSP, pour réduire l’expression de Nanog et GATA6 dans le futur endoderme primitif (Kang et al., 2017). La suppression de la signalisation FGF par l’inhibiteur de MEK PD0325901 (PD032) ou l’inhibiteur de FGFR PD173074 induit une conversion en épiblaste et aucune spécification en endoderme primitif n’est alors possible (McLean et al., 2014 ; Bessonnard et al., 2017). Conformément à la réduction de l’activité MEK pour établir la pluripotence naïve dans l’épiblaste, PD032 est un composant essentiel du cocktail pour dériver et maintenir des cellules souches embryonnaires de souris (Nichols et al., 2009).
Comme chez la souris, la masse cellulaire interne (MCI) dans le blastocyste humain à 6 jours après la fécondation in vitro (FIV), a montré une colocalisation GATA6 et NANOG avant l’apparition de l’endoderme primitif et de l’épiblaste au jour 7 après la fécondation (Roode et al., 2012). Cependant, le traitement par inhibiteur de MEK ou FGFR ne modifie pas le nombre de cellules NANOG, OCT4 ou GATA4+, ce qui suggère que la signalisation FGF-MEK n’est pas requise dans les embryons humains pour la spécification (Roode et al., 2012). Par conséquent, bien que la ségrégation de l’ICM en endoderme primitif et en épiblaste soit similaire chez les deux espèces, les voies de signalisation sont différentes. Ces données suggèrent que des mécanismes moléculaires et cellulaires spécifiques induisent l’initiation et le maintien de la pluripotence des épiblastes chez l’homme.
- Adhérences cellule-cellule
- Arabidopsis thaliana
- Axe antéro-postérieur chez la drosophile
- Biomécanique du développement
- Caenorhabditis elegans
- Concepts principaux
- Contrôle de la traduction
- Contrôle de la transcription
- Contrôle génétique et épigénétique
- Croissance du tube pollinique et double fécondation chez les Angiospermes
- Croissance et guidage axonal
- Des modèles animaux moins classiques
- Développement de l’oeil des Vertébrés
- Développement et évolution
- Et l’Humain ?
- Exercices sur l’ovogenèse, la spermatogenèse et la fécondation
- Exercices sur le contrôle de l’expression des gènes
- Exercices sur le développement des bourgeons de membre
- Exercices sur le développement des muscles striés squelettiques
- Exercices sur le développement des végétaux et les hormones végétales
- Exercices sur les cycles et les divisions cellulaires
- Exercices sur les étapes du développement, les inductions embryonnaires et la mise en place des axes de polarité
- Exercices sur les matrices extracellulaires, le cytosquelette et les adhérences cellule-cellule
- Exercices sur les voies de signalisation
- Glossaire
- Glossaire des termes liés à la génétique
- Glossaire des termes liés au cytosquelette, la matrice extracellulaire, l’adhérence et la migration cellulaire
- Hématopoïèse et développement des cellules du système immunitaire
- Histoire de la biologie cellulaire et de la biologie du développement
- L’acide rétinoïque
- L’apoptose
- L’autophagie
- L’organogenèse
- L’ovogénèse prépare le développement embryonnaire
- La drosophile
- La fécondation
- La formation des somites
- La gastrulation
- La gastrulation (version allégée)
- La métamorphose chez les Hexapodes et les Amphibiens
- La neurogénèse chez les mammifères adultes
- La neurulation
- La poule
- La signalisation calcique
- La souris
- La superfamille TGFβ et ses voies de signalisation
- La voie de signalisation de l’auxine et ses rôles
- La voie de signalisation Hedgehog
- La voie de signalisation Hippo et ses composants YAP/TAZ
- La voie de signalisation Notch
- Le clivage
- Le cytosquelette
- Le destin des cellules et les réseaux de régulation génique
- Le développement des bourgeons de membre
- Le développement des muscles striés squelettiques
- Le développement des organes génitaux et des cellules germinales
- Le développement du cortex
- Le méristème apical caulinaire en phase végétative et lors de la formation d’une fleur
- Le poisson zèbre
- Le xénope
- Les cellules des crêtes neurales
- Les cellules et les gènes en action dans le développement
- Les cellules souches
- Les cellules tumorales
- Les cycles et les divisions cellulaires
- Les étapes du développement
- Les étapes du développement embryonnaire d’Arabidopsis thaliana et leur contrôle
- Les inductions embryonnaires et les gradients de morphogène
- Les matrices extracellulaires animales
- Les organismes modèles
- Les outils pour étudier l’expression et la fonction des gènes
- Les parois des cellules végétales
- Les techniques et les outils pour la biologie cellulaire
- Les transitions épithélio-mésenchymateuses et les migrations cellulaires
- Les vésicules extracellulaires
- Les voies de signalisation
- Mise en place des axes chez les Vertébrés
- Structures et processus cellulaires
- Voies de signalisation WNT
