
Par Patrick Pla, Université Paris-Saclay
La souris domestique (Mus musculus) est un excellent modèle mammifère pour étudier une grande variété de caractères et de maladies, notamment ceux impliqués dans le développement. Néanmoins, il ne faut pas oublier que le dernier ancêtre commun entre la souris et l’Homme a vécu il y a environ 75 millions d’années, ce qui implique que ce qui est découvert chez la souris ne s’appliquera pas forcément à l’Homme.
L’organisation du génome a fortement divergé entre l’homme et la souris depuis leur ancêtre commun. Environ 180 cassures et réassemblages se sont produits dans les lignées humaine et murine. Bien que le nombre de chromosomes soit similaire chez les deux espèces (23 par génome haploïde
chez l’homme contre 20 chez la souris), leurs structures globales diffèrent considérablement. Néanmoins, même après ce remaniement génomique important, il existe de nombreux grands blocs d’ADN dans lesquels l’ordre des gènes est le même chez l’homme et la souris. Ces régions définissent des synténies.

L’introduction de la souris comme modèle en génétique date du début du XXème siècle, à la même époque que pour la drosophile. Dans une série d’articles de 1902 à 1905, Lucien Cuénot a utilisé des souris pour démontrer les lois de Mendel pour la première fois chez les mammifères. À peu près à la même période, William E Castle, un pionnier de l’utilisation de la drosophile pour étudier la génétique, a également commencé à étudier l’hérédité en utilisant comme phénotype la couleur du pelage chez la souris. Abbie Lathrop, institutrice à la retraite à Granby (Massachusetts, États-Unis) avait commencé quelques années plus tôt à élever des souris comme animaux de compagnie, en utilisant des souris achetées à d’autres amateurs et à sélectionner des couleurs de pelage particulières.
Pour faire un exercice de génétique (corrigé) sur la couleur du pelage de la souris, suivre ce lien.
Le laboratoire de Castle ainsi que d’autres ont lancé des programmes de recherche axés sur la génétique de la souris et ont rapidement réalisé la nécessité de créer des souches consanguines. En 1909, la première souche consanguine, DBA, a été créée et l’ère de la génétique de la souris moderne a commencé. Depuis lors, des centaines de souches consanguines ont été développées.
En 1929, CC Little a fondé le Roscoe B Jackson Memorial Laboratory, qui possède désormais le plus grand choix de lignées de souris au monde (plus de 8000 !) Parmi les autres fournisseurs importants figurent Charles River Laboratories, Harlan Laboratories et Taconic Farms, Inc., entre autres. Des lignées d’origine « naturelles » sont à la disposition des chercheurs mais aussi et surtout des lignées générées par différentes techniques de transgénèse et de mutations dirigées (voir plus loin).

Vidéo sur la dissection de l’appareil génital de la souris.

Frise chronologique du développement des souris nouveaux-nés.
La souris est un excellent modèle pour étudier les étapes précoces du développement (clivage) et les étapes post-gastrulation.



Pour la période juste après l’implantation correspondant à la gastrulation, les embryons ont une taille et une topologie qui les rend difficiles à étudier dans l’utérus. Des techniques récentes de culture d’embryons in vitro ont permis de faire se développer des embryons de souris jusqu’à 11 jours après fécondation soit jusqu’à l’organogenèse ! (Aguilera-Castrejon et al., 2021).
Transgénèse chez la souris
Par traitement hormonal, on fait superovuler des souris femelles puis on les accouple avec un mâle. Les zygotes sont rapidement récupérés dans les voies génitales puis incubés in vitro. L’ADN d’intérêt, souvent un gène sous le contrôle d’un promoteur spécifique est injecté dans le pronucléus mâle. Puis on sélectionne les embryons qui ont poursuivi correctement leur développement et on les injecte dans l’utérus d’une femelle pseudo-gestante (la copulation provoque des stimuli mécaniques nécessaires au bon développement de l’utérus pour la gestation alors la femelle est préalablement accouplée avec un mâle vasectomisé). Les souriceaux nés doivent ensuite être sélectionnés pour la présence et l’expression du transgène. En effet, l’insertion du transgène dans le génome ne réussit pas à chaque fois et le transgène peut aussi très bien s’être inséré dans de l’hétérochromatine silencieuse. On effectue une RT-PCR ou alors un test qui permet de révéler l’expression d’un gène rapporteur s’il est présent dans le transgène (par exemple, coloration X-gal si on a mis le gène de la β-galactosidase). De toute manière, il faut toujours étudier plusieurs lignées transgéniques car l’ADN exogène s’insérant n’importe où au hasard dans le génome, il faut vérifier que le phénotype observé n’est pas provoqué par la mutation créée par le point d’insertion mais bien par l’information du transgène lui-même.
Exemple d’application :

Barre d’échelle = 1 mm.
Une version commentée de cette figure est disponible en vidéo.
Source : https://rupress.org/jcb/article/138/6/1343/716/Tissue-specific-Expression-of-the-L1-Cell-Adhesion
Technique de knock-out et CRISPR/Cas9


Le système Cre-Lox
Les knock-out abolissent la fonctionnalité d’un gène depuis le début de son expression. Or parfois un gène peut avoir des fonctions à différents moments du développement. S’il a un rôle vital à une phase précoce, un knock-out ne permettra pas de connaître sa fonction à des phases tardives. Le système Cre-Lox a permis de franchir cet obstacle en rendant possible une délétion d’un gène contrôlée spatio-temporellement au cours du développement. La Cre est une recombinase du bactériophage P1 qui est capable d’exciser toute séquence située entre deux séquences LoxP. Ainsi, il suffit de faire une souris transgénique exprimant Cre sous le contrôle d’un promoteur spécifique ou exprimant une forme activable de Cre (fusion Cre-récepteur aux œstrogènes activable par le tamoxifène) et de la croiser avec une souris où on a introduit par knock-in deux séquences LoxP entourant l’ADN à déléter.
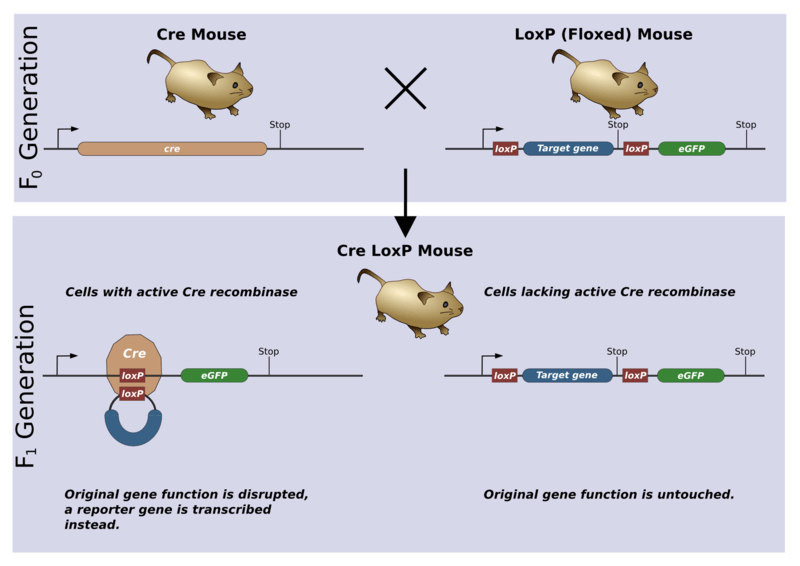
Une version commentée en détails de cette figure est disponible en vidéo. Source : https://en.wikipedia.org/wiki/Cre-Lox_recombination#/media/File:CreLoxP_experiment.png
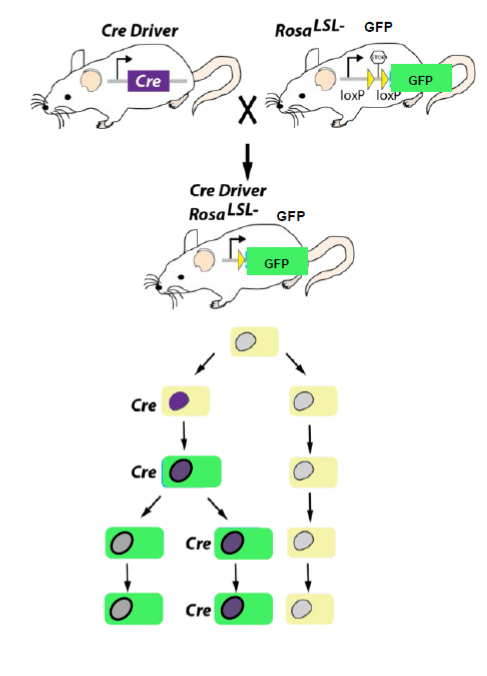
Le système Cre-Lox peut aussi être utilisé pour faire du suivi de lignage cellulaire. L’activation d’un promoteur donné dirige l’expression de la Cre qui va réaliser la délétion d’une séquence générant un codon STOP qui empêche la production d’une protéine rapportrice fonctionnelle. Toutes les cellules qui ont activé le promoteur et aussi ses descendantes vont alors exprimer la protéine rapportrice. Cela est valable y compris pour les cellules où l’activité du promoteur se sera éteinte car la délétion de la séquence générant un codon STOP est définitive et donc une expression transitoire de la Cre suffit.
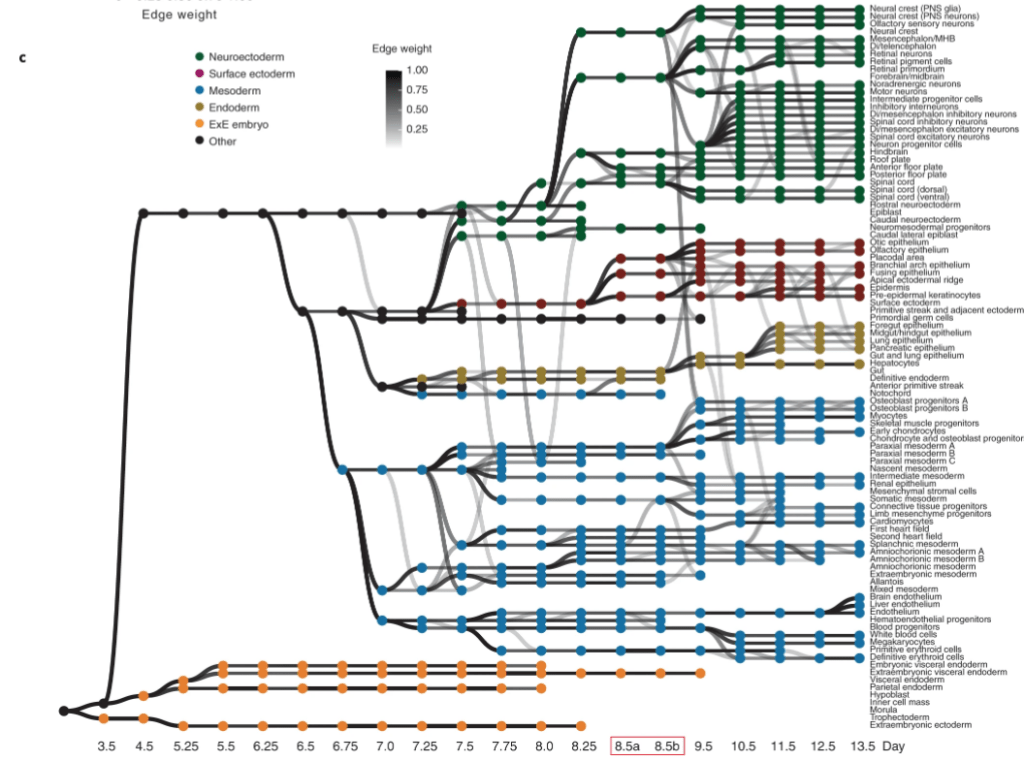
De plus en plus d’études de transcriptomique
Le développement chez la souris, comme dans d’autres modèles, est de plus en plus étudié à l’échelle transcriptomique et cellulaire par scRNAseq (analyse transcriptomique sur cellules isolées). Des lignages basés sur la compilation de plusieurs études à différents stades ont pu être établis (Qiu et al., 2022).
SITES DE RÉFÉRENCE : The Jackson Laboratory et ses plus de 8000 lignées de souris.
Atlas des stades de l’embryogenèse de la souris
AUTRES RESSOURCES :
QUELQUES LABORATOIRES FRANCOPHONES TRAVAILLANT AVEC CE MODELE :
Centre d’études et de recherches thérapeutiques en ophtalmologie (CERTO) – Rétina France, Saclay
Equipe « Reprogrammation épigénétique et Développement » – IGM, Montpellier
Equipe « Mécanismes moléculaires de la différenciation cellulaire dans l’embryon précoce de souris » – GRED, Clermont-Ferrand
Equipe « Signalisation Notch dans les cellules souches et les tumeurs » – Institut Curie, Paris
Equipe « Mécanique du développement des Mammifères » – Institut Curie, Paris
Equipe « Signalisation et progression tumorale » – Institut Curie, Orsay
Equipe « Signalisation, développement et tumeurs cérébrales » – Institut Curie, Orsay
Equipe « Rôle des cils dans le développement et la pathologie du cerveau » – IBENS, Paris
Equipe « Cellules souches et développement musculaire » – Institut Pasteur, Paris
- Adhérences cellule-cellule
- Arabidopsis thaliana
- Axe antéro-postérieur chez la drosophile
- Biomécanique du développement
- Caenorhabditis elegans
- Concepts principaux
- Contrôle de la traduction
- Contrôle de la transcription
- Contrôle génétique et épigénétique
- Croissance du tube pollinique et double fécondation chez les Angiospermes
- Croissance et guidage axonal
- Des modèles animaux moins classiques
- Développement de l’oeil des Vertébrés
- Développement et évolution
- Et l’Humain ?
- Exercices sur l’ovogenèse, la spermatogenèse et la fécondation
- Exercices sur le contrôle de l’expression des gènes
- Exercices sur le développement des bourgeons de membre
- Exercices sur le développement des muscles striés squelettiques
- Exercices sur le développement des végétaux et les hormones végétales
- Exercices sur les cycles et les divisions cellulaires
- Exercices sur les étapes du développement, les inductions embryonnaires et la mise en place des axes de polarité
- Exercices sur les matrices extracellulaires, le cytosquelette et les adhérences cellule-cellule
- Exercices sur les voies de signalisation
- Glossaire
- Glossaire des termes liés à la génétique
- Glossaire des termes liés au cytosquelette, la matrice extracellulaire, l’adhérence et la migration cellulaire
- Hématopoïèse et développement des cellules du système immunitaire
- Histoire de la biologie cellulaire et de la biologie du développement
- L’acide rétinoïque
- L’apoptose
- L’autophagie
- L’organogenèse
- L’ovogénèse prépare le développement embryonnaire
- La drosophile
- La fécondation
- La formation des somites
- La gastrulation
- La gastrulation (version allégée)
- La métamorphose chez les Hexapodes et les Amphibiens
- La neurogénèse chez les mammifères adultes
- La neurulation
- La poule
- La signalisation calcique
- La superfamille TGFβ et ses voies de signalisation
- La voie de signalisation de l’auxine et ses rôles
- La voie de signalisation Hedgehog
- La voie de signalisation Hippo et ses composants YAP/TAZ
- La voie de signalisation Notch
- Le clivage
- Le cytosquelette
- Le destin des cellules et les réseaux de régulation génique
- Le développement des bourgeons de membre
- Le développement des muscles striés squelettiques
- Le développement des organes génitaux et des cellules germinales
- Le développement du cortex
- Le méristème apical caulinaire en phase végétative et lors de la formation d’une fleur
- Le poisson zèbre
- Le xénope
- Les cellules des crêtes neurales
- Les cellules et les gènes en action dans le développement
- Les cellules souches
- Les cellules tumorales
- Les cycles et les divisions cellulaires
- Les étapes du développement
- Les étapes du développement embryonnaire d’Arabidopsis thaliana et leur contrôle
- Les inductions embryonnaires et les gradients de morphogène
- Les matrices extracellulaires animales
- Les organismes modèles
- Les outils pour étudier l’expression et la fonction des gènes
- Les parois des cellules végétales
- Les techniques et les outils pour la biologie cellulaire
- Les transitions épithélio-mésenchymateuses et les migrations cellulaires
- Les vésicules extracellulaires
- Les voies de signalisation
- Les voies de signalisation FGF
- Mise en place des axes chez les Vertébrés
- Structures et processus cellulaires
- Voies de signalisation WNT
