
Par Patrick Pla, Université Paris-Saclay
Il s’agit de la première étape du développement embryonnaire où le zygote puis l’embryon subit des mitoses successives, ce qui divise le cytoplasme issu de l’ovocyte. C’est durant cette période que les cellules germinales (qui forment les gamètes) sont séparées des cellules somatiques, qui sont spécifiées en trois feuillets : ectoderme, mésoderme et endoderme.
- Clivage en spirale
- Clivage radiaire
- Le cas particulier de la drosophile
- Le clivage chez C. elegans
- Compétition cellulaire
Clivage en spirale
Le clivage en spirale est caractérisé par une rotation de 45° du fuseau mitotique par rapport à l’axe animal-végétatif dans la transition du stade quatre à huit cellules.

Cette rotation persiste dans les divisions ultérieures, avec à chaque fois une alternance de sens, soit dextre soit senestre. Finalement, cela se traduit par des cellules situées au pôle animal de l’embryon affichant un arrangement compact en forme de spirale, d’où le nom de ce type de clivage.
Le clivage en spirale est présent chez au moins huit grands groupes d’animaux, incluant les Annélides, les Mollusques et les Plathelminthes (Hejnol, 2010; Henry, 2014). Souvent considéré à tort comme le modèle de clivage typique des Protostomiens, le clivage en spirale est plutôt spécifique et probablement une synapomorphie (caractère dérivé partagé exclusif) des Spiralia. Les Spiralia sont un groupe morphologiquement et écologiquement diversifié comprenant environ 10% des espèces animales connues (Brusca et al., 2016).
La domination de systèmes modèles de Protostomiens qui n’ont pas de clivage en spirale tels l’insecte Drosophila melanogaster ou le nématode Caenorhabditis elegans, tous deux appartenant aux Ecdysozoaires (donc hors des Spiralia), a causé un retard dans l’étude du clivage en spirale.
Clivage radiaire
Chez les amphibiens
Chez les amphibiens, le zygote subit une série de mitoses très rapides qui vont le rendre pluricellulaire. L’ensemble du volume de l’ovocyte est cellularisé : on parle de clivage total ou holoblastique. Le premier plan de clivage est méridien et correspond à l’axe pôle animal/pôle végétatif. Le zygote est divisé en deux cellules de taille similaire, appelées blastomères. Le second plan de division est également méridien mais perpendiculaire au premier.

Bien que similaires en apparence, ces blastomères ne sont pas identiques. Hans Spemann a montré en 1903 que si on sépare les blastomères, seuls ceux qui ont hérité du croissant gris (une région partiellement pigmentée générée par la rotation corticale à la suite de la fécondation) donne des embryons normaux tandis que les autres donnent des structures sans axes définis. Le troisième plan de division est perpendiculaire aux deux précédents, parallèle à l’ »équateur » mais légèrement décalé dans l’hémisphère animal. Les cellules générées n’ont plus les mêmes tailles avec 4 cellules plus petites (appelées micromères) autour du pôle animal et 4 cellules plus grosses (appelées macromères) du côté du pôle végétatif. Puis de nouveaux clivages permettent d’arriver au stade 16 cellules, le stade morula.
Avec de nouvelles divisions, on arrive au stade blastula. Les macromères autour du pôle végétatif sont toujours plus gros (car plus riches en vitellus) que les micromères autour de pôle animal. La blastula se creuse d’une cavité remplie de liquide, le blastocœle, par transport polarisé d’ions (l’eau suit par osmose).

Nieuwkoop a montré en 1969 qu’une blastula d’Amphibien pouvait être divisée en 3 régions :
- Les cellules de la calotte animale autour du pôle animal qui, isolées du reste des cellules, donnent de l’épiderme cilié.
- Les cellules de la zone marginale qui, isolées du reste des cellules, donnent du mésoderme
- Les cellules autour du pôle végétatif qui, isolées du reste des cellules, donnent de l’endoderme.
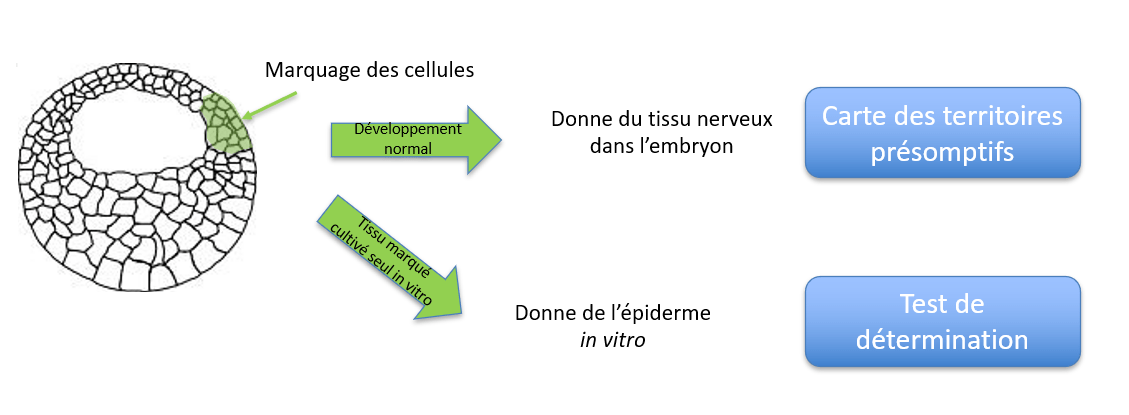
Pendant et à l’issue du clivage, on peut marquer spécifiquement des cellules de l’embryon, laisser la suite du développement se dérouler et observer en quoi ces cellules marquées se développeront plus tard. On établit ainsi une carte des territoires présomptifs. Pour ce faire, on réalise des injections de marqueurs cytoplasmiques fluorescents comme la fluorescéine ou la rhodamine couplée au dextran. Le dextran est un polymère glucidique non dégradable dans le cytoplasme et trop gros pour passer d’une cellule à l’autre ou traverser la membrane plasmique. Il est donc un bon outil pour le suivi du lignage des cellules.

(B) Coupe transversale d’un embryon en fin de neurulation montrant les tissus dérivés du blastomère C1 marqué. Les descendants de C1 contribuent principalement à la formation de la chorde (ou notochorde) et des somites, structures dérivées du mésoderme axial et paraxial. Source : Biologie du développement, Olivier Pourquié, Hermann, 2002

Si on constate un décalage entre le devenir des cellules cultivées isolément et la carte des territoires présomptifs (où les cellules marquées et suivies restent à leur place habituelle dans l’embryon) cela veut dire que les cellules ne sont pas encore déterminées. Par exemple, tout un côté (dorsal) de la calotte animale donne le système nerveux dans les cartes des territoires présomptifs alors que la calotte animale prélevée à différents stades du clivage et cultivée isolée ne donne que de l’épiderme. Cela veut dire qu’au stade où la calotte animale a été prélevée, les signaux inducteurs du système nerveux en provenance de l’extérieur de la calotte animale n’avaient pas encore été reçus.
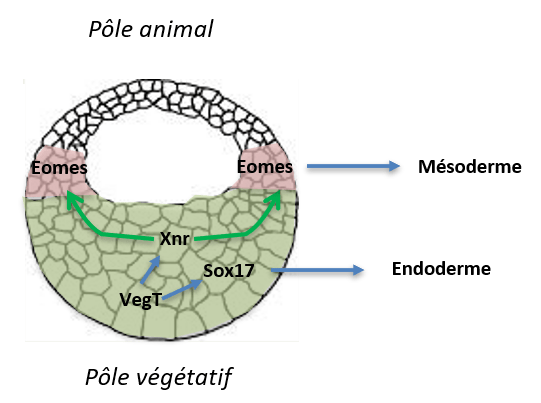
La détermination du mésoderme a lieu pendant la période de clivage :

Des expériences de recombinaisons telles que celles effectuées par Nieuwkoop ont montré que le mésoderme est induit à partir de l’ectoderme à partir de signaux provenant de l’endoderme. On parle d’induction du mésoderme. Ces signaux sont déjà polarisés puisque de l’endoderme dorsal induit du mésoderme dorsal, et de l’endoderme ventral induit du mésoderme ventral. On connait maintenant la signalisation impliquée. Il s’agit de la signalisation Nodal de la famille des ligands TGFβ (ligands Xnr chez le xénope).
L’expression de ces ligands Xnr est activée par VegT, un facteur de transcription dont les ARNm ont été stockés au pôle végétatif lors de l’ovogenèse mais qui n’ont pas été déplacés lors de la rotation corticale. VegT active l’expression de Sox17 qui détermine les cellules de l’hémisphère végétatif en endoderme et il active aussi l’expression des Xnr qui diffusent et induisent le mésoderme dans les cellules sus-jacentes (Skirkanich et al., 2011). Ces gènes sont induits avant même l’activation plus générale de la transcription lors de la MBT (voir plus loin).
Chez les amphibiens, les premières divisions sont très rapides (toutes les 30-35 minutes à 25°C), ce qui est exceptionnel pour une cellule eucaryote : c’est une vitesse qui n’a rien à envier à la prolifération des bactéries ! Le cycle cellulaire est fortement modifié avec une succession de phase S et de phase M, sans phases G1, ni G2. Cela ne laisse absolument pas de temps à une quelconque transcription de se faire.
Lors de la fécondation chez le xénope, le taux de synthèse des protéines augmente fortement et pendant le clivage, un grand nombre de nouvelles protéines sont synthétisées, comme l’ont montré des études de protéomique. Toutes ces protéines sont synthétisées par traduction d’ARNm maternels préformés. Il y a très peu de nouveaux ARN (ARNm, ARNr et ARNt) synthétisés jusqu’au 12ème cycle cellulaire, où l’embryon est composé de 4096 cellules (=212 cellules). A cette étape qui s’appelle la transition mi-blastuléenne (MBT), le cycle cellulaire ralentit permettant à une phase G1 et G2 de se mettre en place. La transcription est activée. Les gènes paternels sont donc transcrits pour la première fois à cette période et avant, c’était le génotype maternel qui contrôle le développement. A ce moment, les divisions deviennent également asynchrones, certains groupes de cellules proliférant plus vite que d’autres.

Qu’est-ce qui déclenche la MBT ? Il semble que ce ne soit pas l’accumulation des divisions cellulaires. Les interactions cellule-cellule ne sont pas non plus impliquées, car des blastomères dissociés subissent la transition en même temps que les embryons intacts. Le facteur clé déclenchant la MBT semble être le ratio ADN/cytoplasme, à savoir la quantité d’ADN présente par unité de masse de cytoplasme. En effet, l’augmentation artificielle de la quantité d’ADN en laissant plus d’un spermatozoïde entrer dans l’ovocyte ou en injectant de l’ADN supplémentaire dans le zygote aboutit à une MBT précoce. Cela suggère qu’il existe une quantité fixe d’un répresseur de transcription présent initialement dans le cytoplasme de l’ovocyte. Comme ce volume est clivé, la quantité de cytoplasme n’augmente pas, mais la quantité d’ADN si (car elle est doublée à chaque réplication). Donc la quantité de répresseur par rapport à l’ADN devient de plus en plus réduite, jusqu’à ce qu’elle soit insuffisante pour se lier à tous les sites disponibles sur l’ADN et la répression est alors levée.
Chez les oiseaux
Chez les oiseaux tels que la poule, le clivage ne concerne qu’une toute petite région du volume de l’ovocyte, le reste restant occupé par le vitellus et restant acellulaire. La première mitose a lieu environ 4 heures après la fécondation et les 16 premières cellules ne sont pas complètement entourées par une membrane plasmique et restent « ouvertes » sur le vitellus. Ensuite, les nouvelles cellules produites sont complètement « fermées ». Des ions et de l’eau sont pompés à partir du blanc d’œuf et s’accumulent dans une cavité formant le blastocoele, sous le disque aplati formé par l’embryon lui-même, appelé blastoderme.

Les axes de l’embryon de poule sont mis en place à ce stade. L’axe dorso-ventral est défini par la proximité avec le vitellus (le côté ventral est le plus proche du vitellus). L’embryon passe alors 20h dans la partie de l’appareil génital appelé utérus et où se dépose la coquille calcaire. La paroi musculeuse de l’utérus provoque une lente rotation de l’œuf (10 à 12 tours par heure) assurant ainsi la distribution homogène des cristaux. Cette rotation a aussi un rôle dans l’établissement de l’axe antéro-postérieur de l’embryon.
Lorsque l’œuf est pondu, l’embryon a terminé son clivage et possède 20.000 à 30.000 cellules. Un sous-ensemble de ces cellules forme un disque quasi-épithélial épais d’une seule couche, l’épiblaste. À la périphérie de l’embryon, l’épiblaste repose sur une couche rigide de plusieurs épaisseurs de grandes cellules mésenchymateuses, qui entrent directement en contact avec le vitellus sous-jacent. Cette portion externe de l’embryon en forme de couronne est connue sous le nom d’Area Opaca (AO). Elle paraît sombre à cause de très nombreuses gouttelettes lipidiques dans les cellules collées contre le vitellus. Dans la partie centrale de l’embryon qui paraît nettement plus claire, l’Area Pellucida (AP), des amas de quelques petites cellules arrondies s’attachent à la face ventrale (inférieure) de l’épiblaste, formant l’hypoblaste primaire. Au cours du développement initial, l’hypoblaste primaire s’aplatit pour former une couche épithéliale de grandes cellules minces, l’hypoblaste. Les cellules épiblastiques AP donnent naissance à l’embryon proprement dit, tandis que l’hypoblaste et l’AO forment des structures extra-embryonnaires.
Durant cette phase, les capacités de régulation de l’embryon de poulet sont remarquables. On peut diviser le disque embryonnaire en 8 et les 8 portions peuvent produire un embryon entier (Sprat et Haas, 1960).
Chez les Mammifères
Le clivage a lieu dans les voies génitales femelles, avant l’implantation. Contrairement aux embryons de drosophile ou de xénope, les premières divisions cellulaires de l’embryon des Mammifères sont très lentes et elles deviennent assez précocement asynchrones.

L’activation du génome zygotique a lieu dans un embryon avec peu de cellules contrairement à la drosophile et au xénope. Cette activation comprend deux vagues : une mineure et une majeure. Chez la souris, la vague mineure a lieu à la fin du stade zygote, tandis que la vague majeure se produit au stade 2 cellules (Schulz et Harrison, 2019). Chez l’homme, la vague mineure a lieu au stade 4 cellules tandis que la majeure se produit au stade 8 cellules. La vague mineure tant chez la souris que chez l’homme est sous le contrôle du facteur de transcription DUX4 (De Iaco et al., 2017). Chez la souris, l’ARN polymérase II qui synthétise les ARNm est positionnée sur de nombreux sites de la chromatine dès le stade zygote mais change de positionnement au stade 2 cellules juste avant la vague majeure. Ce changement de répartition essentiel à l’activation du génome zygotique est sous le contrôle des protéines OBOX (Ji et al., 2023).
Initialement, les blastomères au stade 8 cellules sont lâchement attachés les uns aux autres mais lors de la compaction, l’adhérence cellule-cellule devient nettement plus forte (elle a lieu au stade 8 cellules chez la souris et de manière asynchrone entre les stades 8 et 16 cellules chez l’Homme).


La compaction nécessite la présence d’ions Ca2+ extracellulaires ce qui suggère que des cadhérines, des molécules transmembranaires jouant un rôle dans l’adhérence cellulaire et dont l’activité dépend des ions Ca2+, sont impliqués. Des études plus poussées ont montré que la compaction est causée par l’exocytose de vésicules intracellulaires contenant de la E-cadhérine. Des embryons où les deux allèles du gène codant la E-cadhérine ont été délétés réalisent quand même la compaction ce qui montre que la E-cadhérine impliquée à cette étape est d’origine maternelle (Larue et al., 1994). Au cours de cette étape, les cellules acquièrent une polarité apico-basale comme dans un épithélium.
La compaction de l’embryon humain est également contrôlée par la contractilité cellulaire dépendante des interactions actine-myosine générant une augmentation de la tension superficielle à l’interface cellule-milieu. Cette tension est augmentée 4 fois dans les embryons humains lors de la compaction (et que 2 fois chez la souris) (Turlier et Maître, 2017).
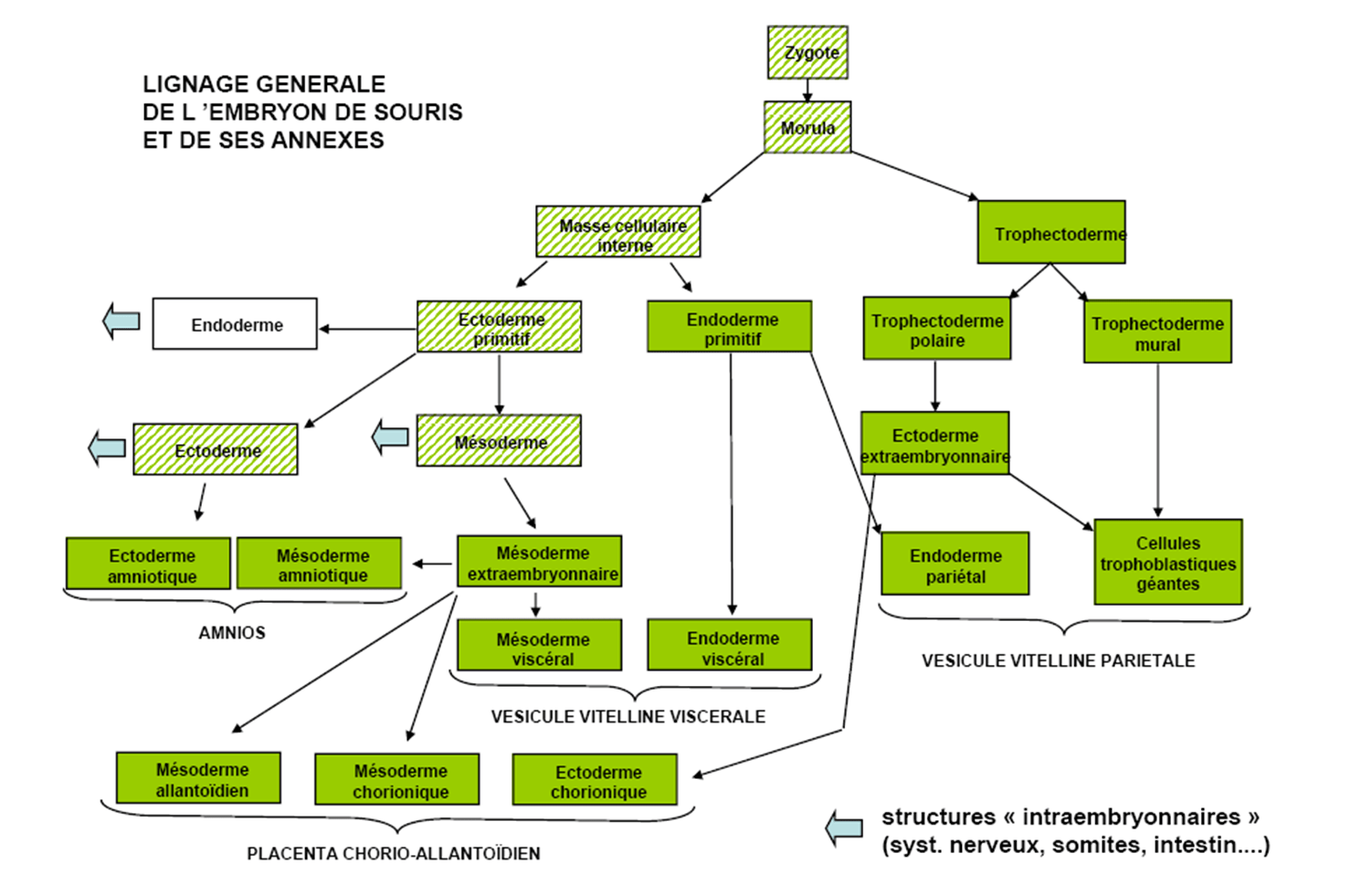
La formation du blastocyste coïncide avec la démarcation des deux premières lignées dans l’embryon préimplantatoire de mammifère (à E3,5 chez la souris) :
- la masse cellulaire interne (MCI) formée d’une quinzaine de cellules seulement et qui donne naissance à l’embryon proprement dit (avec ses trois feuillets), et aussi à l’endoderme et au mésoderme extra-embryonnaires : il s’agit de cellules pluripotentes.
- le trophectoderme (TE) qui génère le placenta.
La spécification de ces lignages est initiée lorsqu’un groupe de cellules est dirigé vers l’intérieur de l’embryon au cours de deux séries de divisions asymétriques aux transitions de 8 à 16 cellules et de 16 à 32 cellules. Les cellules à l’extérieur de l’embryon sont destinées à se différencier en TE extra-embryonnaire, tandis que les cellules à l’intérieur forment la MCI pluripotente. Plus tard, vers E3,5 chez la souris (et vers E5 chez l’Homme), les cellules de la MCI deviendront soit l’épiblaste pluripotent qui donnera naissance au fœtus, soit l’endoderme primitif qui contribuera principalement aux tissus extra-embryonnaires (à ne pas confondre avec l’endoderme de l’embryon qui se formera plus tard).
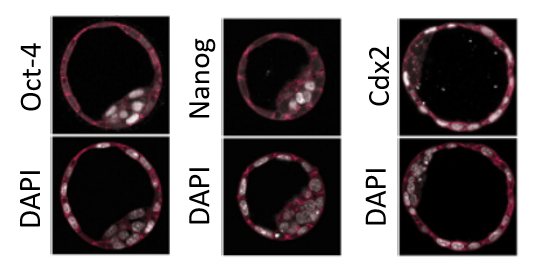
C’est l’activation concomitante des voies de signalisation YAP et Notch qui spécifient les cellules périphériques à se déterminer en trophectoderme, par activation de l’expression de Cdx2 (Watanabe et al., 2017). Dans les cellules de la MCI, YAP reste dans le cytoplasme et ne peut activer l’expression de Cdx2.

Ces données ont été obtenues chez la souris. Chez l’Homme, des expériences suggèrent que CDX2 ne serait pas essentiel pour l’initiation de la formation du trophectoderme mais ce serait GATA3 qui aurait ce rôle (Krendl et al., 2017).
Un transport actif des ions Na+ par le côté basolatéral de l’épithélium formé par le trophectoderme génère un flux osmotique d’eau vers l’intérieur
de l’embryon. De multiples espaces intercellulaires se forment d’abord puis fusionnent en une seule cavité remplie de fluide connue sous le nom de blastocoele (Dumortier et al., 2019).

Les cellules embryonnaires souches (ES) sont dérivées de la masse cellulaire interne (MCI) du blastocyste. Bien qu’il s’agisse d’une population cellulaire transitoire dans l’embryon, les cellules ES cultivées in vitro dans des conditions particulières peuvent subir un auto-renouvellement illimité et sont pluripotentes, donnant naissance à tous les types de cellules (sauf certains dérivés extra-embryonnaires).

D’après https://journals.physiology.org/doi/full/10.1152/physrev.00054.2003
Au stade tardif du blastocyste, trois lignées cellulaires distinctes sont observées : l’épiblaste, l’endoderme primitif, tous deux issus de la MCI et le trophectoderme.


Oct4, Sox2 et Nanog sont des facteurs de transcription qui maintiennent le destin indéterminé des cellules ES et des cellules de l’épiblaste, tandis que le facteur de transcription à homéoboîte Cdx2 régule la mise en place et le maintien de l’identité du trophectoderme (TE) au sein du blastocyste réprimant Oct4 et Nanog dans le TE. Signalons qu’inversement, Nanog réprime Cdx2 dans l’épiblaste et que les inhibitions croisées Nanog/Cdx2 se font directement : Cdx2 se fixant sur le promoteur de Nanog et Nanog se fixant sur le promoteur de Cdx2 (Chen et al., 2009). Gata3 régule le développement du TE en parallèle de Cdx2, et les deux gènes dépendent d’un troisième gène, Tead4. Gata6 contrôle le développement de l’endoderme primitif, antagonisant Nanog, ce qui permet de le distinguer de l’épiblaste.
A E4,5, les populations Nanog+ (épiblaste) et Gata6+ (endoderme primitif) sont ségrégées par la signalisation FGF4 (Yamanaka et al., 2010). Ainsi, les cellules exprimant le récepteur FGFR1 ainsi que la signalisation autocrine FGF4 s’engagent dans l’épiblaste, tandis que la signalisation paracrine FGF4 via FGFR1 et FGFR2 détermine les cellules de l’endoderme primitif. Les niveaux d’ERK actifs relativement faibles dans les cellules FGFR1+ déclenchent une rétroaction négative avec ERK, augmentant l’expression de Nanog et l’identité pluripotente de l’épiblaste. En revanche, la signalisation FGF dans les cellules FGFR1+/FGFR2+ avec des niveaux élevés d’ERK actifs stimule la transcription de cibles alternatives, telles que DUSP, ce qui réduit l’expression de Nanog et active celle de GATA6 dans le futur endoderme primitif (Kang et al., 2017). La suppression de l’activation de ERK par l’inhibiteur PD0325901 (PD032) ou l’inhibiteur de FGFR PD173074 tout comme les mutants perte-de-fonction FGF4-/- ou FGFR1-/-;FGFR2-/- induisent une conversion en épiblaste et aucune spécification en endoderme primitif n’est alors possible (McLean et al., 2014 ; Bessonnard et al., 2017).

ERK inhibe l’expression de Nanog en stimulant le recrutement du facteur de transcription ZFP57 sur de la chromatine contenant H3K9me3. ZFP57 contribue à propager cette marque de chromatine inhibitrice de la transcription. L’absence de la triméthylation H3K9me3 sur les séquences régulatrices de Nanog aboutit à un défaut de différenciation de l’endoderme primitif (Dubois et al., 2022).
Conformément à la réduction de l’activité ERK pour établir la pluripotence naïve dans l’épiblaste, l’inhibiteur de ERK PD032 est un composant essentiel du cocktail pour dériver et maintenir les cellules souches embryonnaires de souris (Nichols et al., 2009).
Comme chez la souris, la MCI dans le blastocyste humain à 6 jours après la fécondation in vitro (FIV), a montré une colocalisation GATA6 et NANOG avant l’apparition de la PE et de l’épiblaste au jour 7 de la FIV (Roode et al., 2012). Cependant, le traitement par un inhibiteur de ERK ou de FGFR ne modifie pas le nombre de cellules NANOG, OCT4 ou GATA4+, ce qui suggère que la signalisation FGF-ERK n’est pas requise dans les embryons humains pour la spécification de la deuxième lignée (Roode et al., 2012). Par conséquent, bien que la séparation de l’ICM en PE et en épiblaste soit similaire chez les deux espèces, leurs voies de signalisation contrastent. Ces données indiquent que des mécanismes moléculaires et cellulaires spécifiques induisent l’initiation et le maintien de la pluripotence des épiblastes chez l’homme.
Lors de l’implantation, le blastocyste établit des contacts avec le tissu maternel et finit par s’attacher à l’endomètre avant d’y pénétrer. L’attachement et l’adhésion à l’endomètre maternel (à E4.75 chez la souris, au jour 7 post-fécondation chez l’Homme) sont médiés par le trophectoderme. A ce stade de développement, deux sous-populations du trophectoderme peuvent être distinguées : la région en contact avec la MCI située au pôle embryonnaire de l’embryon devient le trophectoderme polaire (qui formera l’ectoderme extra-embryonnaire qui contribue fortement à la formation du placenta) et la région entourant la cavité du blastocœle localisée au pôle abembryonnaire devient le trophectoderme mural (qui formera la couche externe du vésicule vitelline pariétale).
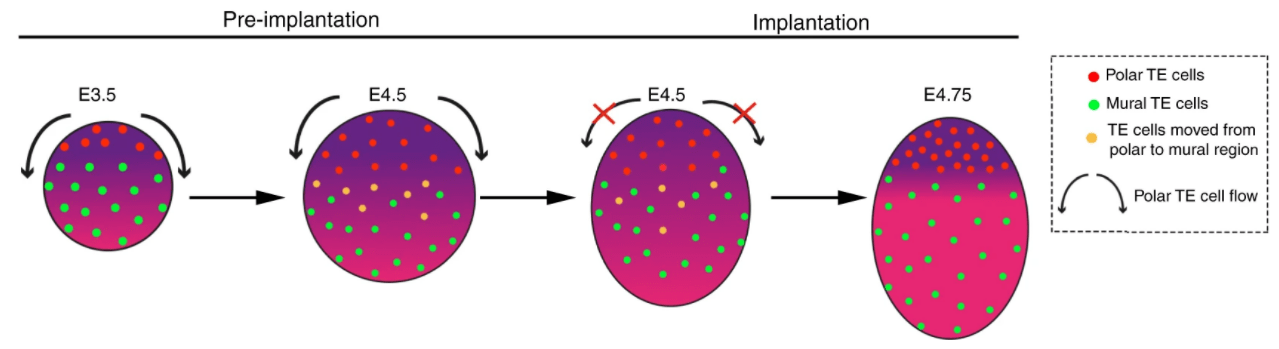
Les cellules du trophectoderme mural perdent l’expression de Cdx2 contrairement aux cellules du trophectoderme polaire. Malgré une potentialité similaire au début du stade blastocyste, ils finissent par se différencier à cause de l’exposition différentielle à la signalisation inductive émanant de la MCI. Les cellules du trophectoderme polaire subissent une transition épithélio-mésenchymateuse et prennent un caractère invasif. Au contact avec l’endomètre utérin maternel, le trophectoderme polaire prolifère et se différencie en cytotrophoblaste et en syncytiotrophoblaste à cellules multinucléés qui dirigent la pénétration du blastocyste à l’intérieur de l’endomètre.


La différenciation des cellules en syncytiotrophoblaste dépend de l’expression du facteur de transcription TBX3.

Les grossesses extra-utérines surviennent lorsqu’un ovule fécondé s’implante et se développe à l’extérieur de l’utérus. Dans presque tous les cas, cela se produit dans la trompe de Fallope, entraînant une grossesse tubaire. Le contrôle du mouvement des embryons dans l’oviducte et vers l’utérus durant le clivage joue donc un rôle essentiel. Le transit des embryons à travers l’oviducte est dû à l’action combinée de battements des cils de l’épithélium de l’oviducte dans un sens précis, ses sécrétions régulées et les contractions périodiques des muscles environnants (Bianchi et al., 2021).
Dans la période précédant l’implantation et pendant l’implantation, l’endomètre utérin est loin d’être passif. Les glandes utérines nourrissent bien sûr l’embryon et sécrètent également du LIF (pour Leukemia Inhibitory Factor) qui déclenche dans les cellules du trophectoderme une cascade de signalisation qui est indispensable à une bonne implantation (Kelleher et al., 2018).

C’est durant le clivage qu’a lieu l’inactivation d’un des chromosomes X chez les femelles des Mammifères. Cette inactivation permet de compenser le fait que les femelles XX ont 2 allèles par gène du chromosome X alors que les mâles XY n’en ont qu’un seul. C’est de manière aléatoire dans chaque cellule que l’inactivation concerne soit le chromosome X d’origine paternel soit celui d’origine maternel. Les femelles des Mammifères sont donc des mosaïques génétiques en ce qui concerne les gènes portés par le chromosome X.

Le chromosome inactivé forme une structure d’hétérochromatine dense appelée corps de Barr. Cette condensation est sous le contrôle d’un long ARN non codant (lncRNA) de 17 kb appelé Xist et qui est transcrit seulement à partir du chromosome X qui sera inactivé. Cet ARN recrute plusieurs facteurs qui modifient la structure chromatinienne de ce chromosome et le réduisent au silence (Boeren et Gribnau, 2021). Tant que les cellules sont pluripotentes durant le clivage, la transcription de Xist est bloquée directement par Nanog, Oct4 (Pou5f1) et Sox2 (Minkovsky et al., 2012). Un autre ARN, Tsix qui est anti-sens de l’ARN Xist, est transcrit à partir du chromosome X qui reste actif et le protège contre l’action de Xist (Lee et Lu, 1999).
Le cas particulier de la drosophile
Après la fécondation et la fusion des pronucléi mâle et femelle, le noyau zygotique subit 13 cycles de réplication en l’absence de division cellulaire ce qui génère un embryon unicellulaire avec environ 5000 noyaux (Schmidt et Grosshans, 2018).

{kind=link}

Cette figure illustre les différentes phases du clivage précoce de l’embryon de drosophile, caractérisé par une division nucléaire sans cytodiérèse, conduisant à la formation d’un blastoderme syncytial puis cellulaire :
A. Cycles 4–6 : Les noyaux issus des divisions synchrones restent regroupés au centre de l’embryon.
B. Cycles 8–9 : Migration des noyaux vers la périphérie (cortex) de l’embryon.
C. Cycles 10–13 (Blastoderme syncytial) : Les noyaux s’alignent en périphérie, créant une couche syncytiale sans membranes cellulaires. Les cellules germinales (cellules polaires) commencent à se former à l’arrière.
D. Interphase 14 (Cellularisation) : Chaque noyau périphérique devient entouré par une membrane plasmique, initiant la cellularisation.
E. Blastoderme cellulaire : L’embryon est désormais constitué de cellules individuelles entourant le vitellus. L’image en coupe montre une coloration de l’ADN (en rouge), révélant la disposition périphérique des noyaux. Source : https://onlinelibrary.wiley.com/doi/abs/10.1002/bies.10184
Les 9 premiers cycles de réplication/mitose se produisent profondément dans le vitellus de l’embryon et sont très rapides, toutes les 10 minutes. Au cycle nucléaire 10, les noyaux migrent vers la périphérie et commencent à organiser la formation de structures d’actine corticale qui initient ensuite la formation de sillons de type cytodiérèse qui aboutissent à la véritable cellularisation de l’embryon. L’embryon est cellularisé au cycle nucléaire 14. La cellularisation se fait en 10 minutes durant lesquels des vésicules cytoplasmiques fusionnent pour former la membrane plasmique.
Ce moment donne lieu également à la transition mi-blastuléenne qui voit s’activer la transcription de nouveaux ARNm (dits zygotiques) et l’élimination des ARNm maternels. Cependant, signalons que quelques gènes zygotiques sont exprimés à partir du cycle nucléaire 7.

L’activation massive de la transcription au cycle nucléaire 14 s’accompagne de modifications importantes de la chromatine qui est notamment permise par la présence de facteurs dit pionniers tels que le facteur de transcription d’origine maternel Zelda qui se lie aux nucléosomes et rendent l’ADN plus accessible (Melody Foo et al., 2014). La perte de Zelda aboutit à une baisse drastique des gènes dont la transcription est activée lors de la transition mi-blastuléenne.

La transition mi-blastuléenne est aussi permise par la levée de l’inhibition apportée par des protéines comme Tramtrack, Supressor of Hairless (Su[H]) et Runt.
La dégradation des ARNm d’origine maternelle est sous la dépendance de Pumilio, BRAt et Smaug mais aussi du cluster de microARN miR-309 qui contient 8 microARN et qui est responsable de la dégradation d’environ 400 ARNm maternels différents ! (Bushati et al., 2008). L’expression de ces microARN est sous la dépendance de Smaug (Benoit et al., 2009).
Le clivage chez C. elegans
Vidéo introductive : clivage chez C. elegans avec le nom des cellules pour le lignage :
Le premier clivage donne naissance à une grande cellule somatique, le blastomère AB, qui donne naissance à la majeure partie de l’hypoderme et du système nerveux et à une plus petite cellule P1, qui, en plus de donner la lignée germinale et les gonades produit également l’intestin et la plupart des muscles de l’animal.

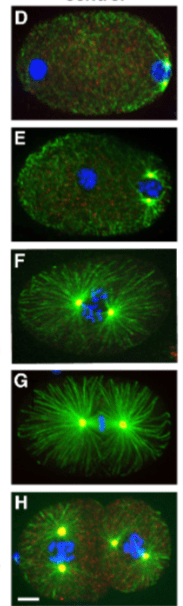

P1 subit trois divisions asymétriques successives, générant à chaque fois un blastomère somatique (EMS, C et D) et un blastomère germinal (P2, P3 et P4).

Les blastomères germinaux diffèrent de leurs sœurs somatiques par leur petite taille, leur progression plus lente dans le cycle cellulaire et leur répression de la transcription. Au stade ∼100 cellules, P4 se divise symétriquement pour générer Z2 et Z3 qui prolifèrent tout au long du développement larvaire et donnent naissance aux spermatozoïdes et aux ovocytes de la lignée germinale adulte.
La structuration initiale de l’embryon de C. elegans est contrôlée par une collection d’ARNm et de protéines codés par la mère qui contrôlent la taille, la position et l’identité des premiers blastomères.
Les sept protéines PAR orchestrent les divisions asymétriques dans l’embryon précoce de C. elegans. Les embryons mutants par ne parviennent pas à diviser les déterminants somatiques et germinaux au cours des premières divisions. Les protéines PAR contrôlent également le positionnement asymétrique du fuseau mitotique qui se traduit par des blastomères plus gros et plus petits. Après leur identification initiale en tant que mutations létales embryonnaires à effet maternel chez C. elegans, il a été démontré que les PAR contrôlent la polarisation d’une grande variété de types de cellules animales qui réalisent une division asymétrique.
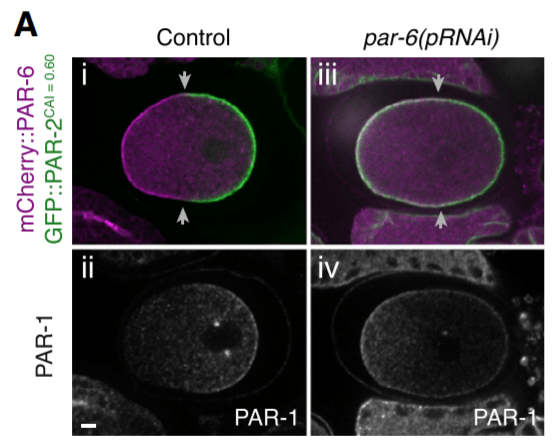
Deux sous-ensembles de protéines PAR se concentrent dans des domaines corticaux antérieurs et postérieurs distincts dans le zygote polarisé de C. elegans. Le cortex antérieur est occupé par les protéines PAR-3 et PAR-6 et la kinase PKC-3 (aPKC chez les mammifères), qui sont collectivement appelées les PAR antérieures. Le domaine cortical postérieur opposé est occupé par la kinase PAR-1 et la protéine PAR-2 (spécifique aux nématodes), appelées PAR postérieures. Deux protéines PAR supplémentaires, la kinase PAR-4 et la protéine PAR-5, sont localisées symétriquement dans le cytoplasme et au niveau du cortex cellulaire.

Les protéines PAR antérieures et postérieures s’associent au cortex cellulaire via une combinaison d’interactions protéine/protéine et une association directe avec des phospholipides. Les expériences de suivi de FRAP avec des protéines fusion GFP::PAR-2 et GFP::PAR-6 démontrent que les protéines PAR sont très dynamiques au niveau du cortex cellulaire, diffusant latéralement le long du cortex et s’échangeant entre le cortex et le cytoplasme.
PKC-3 phosphoryle PAR-1 et PAR-2, les obligeant à se dissocier du cortex et empêchant ainsi leur invasion dans le domaine PAR antérieur. De même, PAR-1 et PAR-2 empêchent les PAR antérieurs d’envahir le domaine postérieur du cortex, en partie par la phosphorylation de PAR-1 et de PAR-3 qui provoque leur dissociation du cortex. La réduction de l’activité de l’un ou l’autre des complexes PAR conduit à la croissance du domaine PAR opposé, ce qui indique que l’équilibre entre les activités PAR antérieures et postérieures aident à positionner leur interface au milieu de l’axe antéro-postérieur.
Avant la polarisation, les protéines PAR sont distribuées symétriquement le long de l’axe antéro-postérieur avec les protéines PAR antérieures enrichies au niveau du cortex cellulaire et les PAR postérieures dans le cytoplasme. La rupture de la symétrie provient de la fécondation. Le centrosome du spermatozoïde, qui est associé au pronucléus mâle à l’extrémité postérieure du zygote, déclenche deux voies de ruptures de symétrie qui établissent la polarité PAR corticale. Dans la première voie, le centrosome initie le flux d’un réseau d’actomyosine cortical vers l’antérieur qui repousse PAR-6 (et vraisemblablement PAR-3 et PKC-3) de l’arrière vers le cortex antérieur, permettant ainsi à PAR-2 et PAR-1 de se concentrer dans le cortex postérieur. Le centrosome initie également une voie de rupture de symétrie, dépendante de PAR-2, qui peut établir la polarité en absence d’actine et de myosine. PAR-2 se lie aux microtubules émanant du centrosome, le protégeant de la phosphorylation par PKC-3. Cette protection permet à PAR-2 de se concentrer sur une petite région du cortex près du centrosome malgré la présence de PKC-3. Le PAR-2 cortical recrute son partenaire de liaison PAR-1 à partir du cytoplasme et, ensemble, PAR-2 et PAR-1 sont capables de provoquer la dissociation des PAR antérieurs du cortex via la phosphorylation de PAR-3. L’élimination des PAR antérieurs permet à plus de PAR-1 et PAR-2 d’atteindre le cortex, initiant ainsi une boucle de rétroaction positive qui entraîne l’expansion du domaine PAR postérieur et la contraction du domaine PAR antérieur.
Signalons que les protéines PAR jouent un rôle général dans la polarisation des épithéliums. Leur première intervention survient lors de la compaction chez la souris, entre le deuxième et le troisième jour après la fécondation, au stade de 8 cellules. Les protéines Par y définissent des domaines apical et basolatéral définis. Le domaine apical s’enrichit du complexe Par3-Par6-aPKC, tandis que le domaine basolatéral s’enrichit de protéines d’adhésion cellulaire (Plusa et al., 2005).
Compétition cellulaire
Au cours des divisions, il y a une compétition entre les cellules. C’est un phénomène par lequel les cellules d’une population « surpassent » les cellules moins aptes dans le même groupe. Chez la drosophile et la souris, les cellules exprimant des niveaux plus élevés de Myc, connus pour activer l’anabolisme cellulaire, surpassent les cellules avec des niveaux de Myc inférieurs. Ces expériences ont montré que ce n’est pas le niveau absolu d’expression de Myc qui compte mais le fait qu’elle exprime un niveau de Myc plus élevé que sa voisine.
Dans le cas des embryons de souris, les cellules qui perdent la compétition finissent par mourir par apoptose. Les restes de ces cellules sont phagocytés par la cellule voisine «plus en forme» qui a une expression de Myc plus élevée (Claveria et al., 2013; Sancho et al., 2013). Le pic d’apoptose lié à la compétition cellulaire a lieu dans l’épiblaste, avant la spécification des 3 feuillets. Durant cette période, les cellules sont beaucoup plus suscpetibles de mourir par apoptose car il y a une expression plus forte de la protéine pro-apoptotique BIM (Pernaute et al., 2014) et donc la compétition cellulaire est plus drastique. Ce mécanisme d’élimination des cellules est un phénomène naturel sélectionnant les cellules les plus aptes pour contribuer au pool de progéniteurs pluripotents qui formeront les 3 feuillets embryonnaires de l’embryon de souris.
Ces mécanismes sont aussi présents dans les tumeurs et également dans les cultures de cellules embryonnaires souches (ES). Il existe une expression hétérogène de Myc dans des cellules ES de souris en culture. Ce déséquilibre Myc entraîne une compétition cellulaire et nécessite un contact cellulaire direct entre le vainqueur et la cellule éliminée. Des mécanismes biomécaniques via le cytosquelette d’actine sont également à l’oeuvre et le co-facteur de transcription YAP en aval des stiumuli biomécaniques se retrouve phosphorylé puis dégradé dans les cellules qui seront éliminées. En revanche, YAP entre dans le noyau et régule la transcription dans les cellules qui survivent (Price et al., 2021).

L’étape suivante du développement est la gastrulation.
CARTE MENTALE

EN DIRECT DES LABOS :
EQUIPES DE RECHERCHE FRANCOPHONES TRAVAILLANT SUR CE THEME :
Equipe « Embryon et pluripotence : épigénétique et environnement » – INRAE Jouy-en-Josas
Equipe « Différenciation des lignages Cellulaires dans l’Embryon précoce de souris » – iGReD , Clermont-Ferrand
- Adhérences cellule-cellule
- Arabidopsis thaliana
- Axe antéro-postérieur chez la drosophile
- Biomécanique du développement
- Caenorhabditis elegans
- Concepts principaux
- Contrôle de la traduction
- Contrôle de la transcription
- Contrôle génétique et épigénétique
- Croissance du tube pollinique et double fécondation chez les Angiospermes
- Croissance et guidage axonal
- Développement de l’oeil des Vertébrés
- Développement et évolution
- Des modèles animaux moins classiques
- Et l’Humain ?
- Exercices sur l’ovogenèse, la spermatogenèse et la fécondation
- Exercices sur le contrôle de l’expression des gènes
- Exercices sur le développement des bourgeons de membre
- Exercices sur le développement des muscles striés squelettiques
- Exercices sur le développement des végétaux et les hormones végétales
- Exercices sur les étapes du développement, les inductions embryonnaires et la mise en place des axes de polarité
- Exercices sur les cycles et les divisions cellulaires
- Exercices sur les matrices extracellulaires, le cytosquelette et les adhérences cellule-cellule
- Exercices sur les voies de signalisation
- Glossaire
- Glossaire des termes liés au cytosquelette, la matrice extracellulaire, l’adhérence et la migration cellulaire
- Glossaire des termes liés à la génétique
- Hématopoïèse et développement des cellules du système immunitaire
- Histoire de la biologie cellulaire et de la biologie du développement
- L’acide rétinoïque
- L’apoptose
- L’autophagie
- L’organogenèse
- L’ovogénèse prépare le développement embryonnaire
- La drosophile
- La fécondation
- La formation des somites
- La gastrulation
- La gastrulation (version allégée)
- La métamorphose chez les Hexapodes et les Amphibiens
- La neurogénèse chez les mammifères adultes
- La neurulation
- La poule
- La signalisation calcique
- La souris
- La superfamille TGFβ et ses voies de signalisation
- La voie de signalisation de l’auxine et ses rôles
- La voie de signalisation Hedgehog
- La voie de signalisation Hippo et ses composants YAP/TAZ
- La voie de signalisation Notch
- Le cytosquelette
- Le développement des bourgeons de membre
- Le développement des muscles striés squelettiques
- Le développement des organes génitaux et des cellules germinales
- Le développement du cortex
- Le destin des cellules et les réseaux de régulation génique
- Le méristème apical caulinaire en phase végétative et lors de la formation d’une fleur
- Le poisson zèbre
- Le xénope
- Les étapes du développement
- Les étapes du développement embryonnaire d’Arabidopsis thaliana et leur contrôle
- Les cellules des crêtes neurales
- Les cellules et les gènes en action dans le développement
- Les cellules souches
- Les cellules tumorales
- Les cycles et les divisions cellulaires
- Les inductions embryonnaires et les gradients de morphogène
- Les matrices extracellulaires animales
- Les organismes modèles
- Les outils pour étudier l’expression et la fonction des gènes
- Les parois des cellules végétales
- Les techniques et les outils pour la biologie cellulaire
- Les transitions épithélio-mésenchymateuses et les migrations cellulaires
- Les vésicules extracellulaires
- Les voies de signalisation
- Les voies de signalisation FGF
- Mise en place des axes chez les Vertébrés
- Structures et processus cellulaires
- Voies de signalisation WNT