
EXERCICE 1
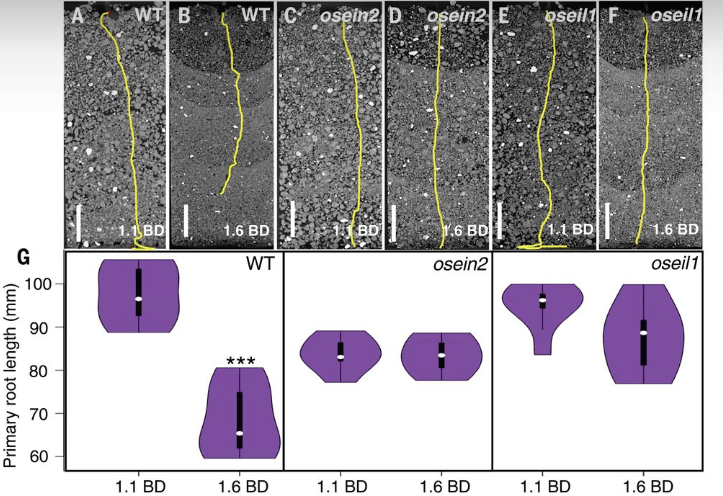
*1.1 Quel est l’effet de la compaction du sol sur la croissance des racines des plants de riz sauvages ?
*1.2 Que mettent en évidence les résultats avec les mutants ?
**1.3 Quelles expériences complémentaires proposeriez-vous pour conforter un modèle de l’effet de la compaction du sol sur les racines ?
**1.4 Quel pourrait être les applications de ce qui mis en évidence ici en agriculture ?
EXERCICE 2
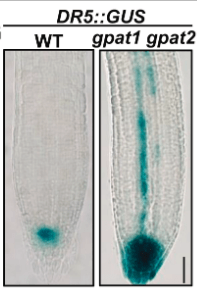
*2.1 Que permettent de voir les plantes transgéniques DR5::GUS ?
*2.2 Analysez le résultat et posez des hypothèses pour l’expliquer.
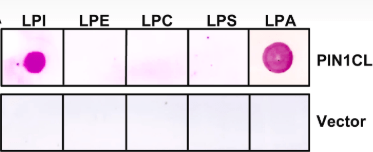
*2.3 Que peut-on déduire de cette expérience ? Interprétez par rapport aux résultats en 2.2
EXERCICE 3
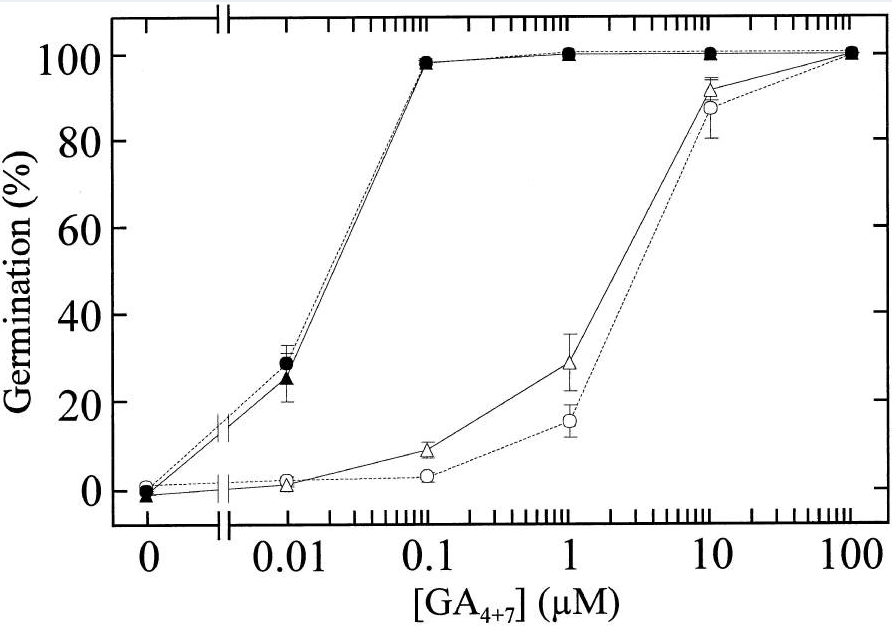
*3.1 Pourquoi ajoute-t-on du tetcyclacis tout en ajoutant des GA exogènes ?
*3.2 Quels sont les effets mis en évidence en comparant les courbes avec les points formant des cercles ?
**3.3 Quelles informations supplémentaires nous apportent les courbes avec les points formant les triangles ?
EXERCICE 4

*4.1 Caractérisez le phénotype du mutant.
**4.2 Trouvez une ou plusieurs hypothèses pour expliquer les origines génétiques du phénotype du mutant.
REPONSES
1.1 Il y a une moindre longueur de la racine primaire qui correspond sans doite à une moindre croissance. 1.2 La compaction du sol n’a plus d’effets sur la croissance de la racine si la voie de transduction de l’éthylène est altérée (à différents niveaux). Cela suggère que c’est l’éthylène qui est responsable de la plus faible croissance des racines dans des sols compactés. 1.3 Il faudrait augmenter les concentrations d’éthylène dans des racines de plants de riz sauvages qui poussent sur des sols peu compactés et on s’attend à observer une moindre croissance de ces racines. 1.4 La compaction des sols est un problème qui limite les rendements agricoles. L’inihibition de la voie de l’éthylène pourrait permettre aux plantes de développer un appareil racinaire normal, même dans des sols compactés.
2.1 Chez les végétaux, on utilise comme gène rapporteur GUS codant la β-glucuronidase dont le substrat est l’acide 5-bromo-4-chloro-3-indolyl-β-D-glucuronique (X-Gluc). Le clivage de X-Gluc sous l’action du GUS produit un précipité bleu. Ici, on suit l’expression du gène rapporteur de l’activité de l’auxine DR5::GUS (GUS est mis sous le contrôle de séquences régulatrices de la transcription activées par la voie de signalisation de l’auxine (DR5)). 2.2 On observe que l’activité GUS qui est sous le contrôle de l’auxune est plus forte et plus étendue, ce qui indique un excès de signalisation provenant de l’auxine. Ainsi donc les deux enzymes GPAT1 et GPAT2 ont un rôle inhibiteur sur la signalisation de l’auxine. Cela peut provenir d’une inhibition de la production d’auxine ou alors d’une inhibition de son transport. Il est possible que ce ne soit pas directement la signalisation liée auxine qui est contrôlée par GPAT1 et GPAT2 mais une autre voie inhibitrice de cette signalisation. 2.3 On détecte la présence de HIS et donc de la boucle cytoplasmique de PIN1 en présence de LPI et de LPA mais pas des autres lysophospholipides (et le contrôle négatif est bien négatif). PIN1 est un transporteur d’auxine, ce qui suggère que GPAT1 et GPAT2 contrôlent le transport de l’auxine via certains des lysophospholipides qu’ils génèrent (LPI et LPA).
3.1 On souhaite s’affranchir de la production de GA endogène par la graine (qui pourrait être variable d’une graine à l’autre) et contrôler précisément la concentration de GA qui agit sur la germination. 3.2 Le GA exogène stimule la germination dans tous les cas mais avec un tégument altéré les doses nécessaires pour arriver à faire germer les graines sont 100 fois plus faibles. Cela peut provenir de ce que le GA exogène pénètre mieux dans les graines avec le tégument altéré mais aussi dû à l’effet inhibiteur d’un tégument intact sur l’action de GA, indépendamment des problèmes de pénétration du GA exogène. 3.3 L’expérience montre que lorsque c’est le père qui porte la mutation tt12, on retrouve la même situation que pour les graines issues de plantes sauvages avec la nécessité de fortes doses de GA pour déclencher la germination, tandis que lorsque c’est la mère qui est porteuse de la mutation, on se retrouve dans la même situation que les graines issues de 2 plantes mutantes car la germination est déclenchée par de faibles doses de GA. Cela indique que ce sont des éléments codés par le génome maternel (sans doute dans les ovaires) qui déterminent l’état altéré ou non du tégument de la graine qui fait changer sa sensibilité aux GA pour le déclenchement de la germination.
4.1 La fleur du mutant ne contient pas de pétales colorées et ne semble pas contenir de pièces fertiles (étamines, carpelles). En revanche, les pièces présentes ressemblent à des sépales avec des longueurs décroissantes au fur et à mesure que l’on se rapproche du sommet. Il s’agit donc d’une mutation homéotique (un élément anatomique à la place d’un autre). 4.2 Selon le modèle ABCE de contrôle génétiques des pièces florales, des fleurs ne comportant que des sépales sont formées en absence d’activité des produits des gènes de type B et C et n’expriment que les gènes de type A (ainsi que les gènes de type E pour que des sépales soient formées et non pas des feuilles). L’absence d’activité des produits des gènes B et C peut provenir de mutations perte-de-fonction dans leurs séquences codantes ou de mutations dans leur séquences régulatrices.
